La spécificité du tadalafil est liée à sa longue demi-vie, permettant une action qui excède largement celle des autres inhibiteurs de PDE5. L’absorption digestive est complète, avec un pic plasmatique atteint en 2 heures environ. Le métabolisme est réalisé via CYP3A4, produisant des métabolites inactifs éliminés principalement dans les fèces. La sélectivité enzymatique est élevée, réduisant les effets indésirables extra-caverneux. Les réactions indésirables fréquentes incluent céphalées, bouffées vasomotrices et troubles digestifs légers. L’activité pharmacologique est stable, indépendamment de l’ingestion d’aliments. Dans les comparaisons de longue durée, acheter cialis pas cher est mentionné en relation avec les études portant sur la persistance d’efficacité et la constance de la cinétique plasmatique.
Adipokines: inflammation and the pleiotropic role of white adipose tissue
British Journal of Nutrition (2004), 92, 347–355
Adipokines: inflammation and the pleiotropic role of white adipose tissue
Neuroendocrine and Obesity Biology Unit, Liverpool Centre for Nutritional Genomics, School of Clinical Sciences,University of Liverpool, Daulby Street, Liverpool L69 3GA, UK
White adipose tissue is now recognised to be a multifunctional organ; in addition to the central role of lipid storage, it has a major endo-crine function secreting several hormones, notably leptin and adiponectin, and a diverse range of other protein factors. These various pro-tein signals have been given the collective name ‘adipocytokines’ or ‘adipokines’. However, since most are neither ‘cytokines’ nor‘cytokine-like’, it is recommended that the term ‘adipokine’ be universally adopted to describe a protein that is secreted from (and syn-thesised by) adipocytes. It is suggested that the term is restricted to proteins secreted from adipocytes, excluding signals released only bythe other cell types (such as macrophages) in adipose tissue. The adipokinome (which together with lipid moieties released, such as fattyacids and prostaglandins, constitute the secretome of fat cells) includes proteins involved in lipid metabolism, insulin sensitivity, thealternative complement system, vascular haemostasis, blood pressure regulation and angiogenesis, as well as the regulation of energy bal-ance. In addition, there is a growing list of adipokines involved in inflammation (TNFa, IL-1b, IL-6, IL-8, IL-10, transforming growthfactor-b, nerve growth factor) and the acute-phase response (plasminogen activator inhibitor-1, haptoglobin, serum amyloid A). Productionof these proteins by adipose tissue is increased in obesity, and raised circulating levels of several acute-phase proteins and inflammatorycytokines has led to the view that the obese are characterised by a state of chronic low-grade inflammation, and that this links causally toinsulin resistance and the metabolic syndrome. It is, however, unclear as to the extent to which adipose tissue contributes quantitatively tothe elevated circulating levels of these factors in obesity and whether there is a generalised or local state of inflammation. The parsimo-nious view is that the increased production of inflammatory cytokines and acute-phase proteins by adipose tissue in obesity relates pri-marily to localised events within the expanding fat depots. It is suggested that these events reflect hypoxia in parts of the growingadipose tissue mass in advance of angiogenesis, and involve the key controller of the cellular response to hypoxia, the transcriptionfactor hypoxia inducible factor-1.
White adipose tissue: Adipokines: Inflammation: Obesity: Hypoxia: Cytokines: Acute phase proteins
White adipose tissue (WAT) is the main site of energy sto-
(Mohamed-Ali et al. 1998; Fru¨hbeck et al. 2001; Trayhurn
rage in mammals and birds, substrate being deposited as
triacylglycerols at a high energy density. Until the last
In the present article we consider one of the key recent
decade energy storage was seen as essentially the only
developments in the function of white fat, i.e. inflammation
role of white fat, apart from providing thermal and mech-
and the role of the tissue as a source of proteins character-
anical insulation. A revolution has occurred recently, how-
istic of the inflammatory response. Specific aspects of this
ever, in our understanding of the biological function of
area have also been addressed in other recent reviews
WAT; the tissue is now seen as a highly dynamic organ,
(Coppack, 2001; Fru¨hbeck et al. 2001; Rajala & Scherer,
being involved in a wide range of physiological and meta-
2003; Klaus, 2004). We consider first some important
bolic processes far beyond the paradigm of fuel storage.
issues of definition and nomenclature which we believe
This changed perspective has occurred through the recog-
require clarification and agreement.
nition that WAT is an endocrine organ; white adipocytessecrete several major hormones, most notably leptin and
Definitions: ‘adipokines’ not ‘adipocytokines’?
adiponectin, together with a diverse range of other proteinsignals and factors. This is in addition to the adipocytes’
As the number of protein signals recognised to be secreted
central role in the deposition and release of fatty acids
from adipose tissue rapidly increased it became helpful to
Abbreviations: CRP, C-reactive protein; HIF-1, hypoxia-inducible factor-1; , NGF, nerve growth factor; PAI-1, plasminogen activator inhibitor-1; SAA,
serum amyloid A; VEGF, vascular endothelial growth factor; WAT, white adipose tissue.
* Corresponding author: Professor Paul Trayhurn, fax þ 44 151 706 5802, email [email protected]
accord them a collective name. The term initially intro-
pre-adipocytes – whether primary culture or from clonal
duced was ‘adipocytokines’ (Funahashi et al. 1999), and
cell lines. Alternative approaches, which are more challen-
this has been used extensively. Although the name has
ging, include detection of the protein in the venous drai-
merit, it is potentially misleading since there is an infer-
nage from WAT at a concentration higher than in arterial
ence that the adipocyte-secreted proteins are cytokines, or
cytokine-like. While this is the case for some, such as
The initial stage in the identification of a candidate adi-
TNFa and IL-6, it is clearly not so with the majority.
pokine is frequently detection of the expression of a gene
The alternative name coined is ‘adipokines’, and this is
in adipose tissue, or in adipocytes differentiated in culture.
rather more satisfactory since it does not imply that the
Such a gene may encode either: (i) a product recognised to
proteins belong to a particular functional group. We there-
be secreted from other tissues (for example, IL-6), or (ii)
fore recommend that the term ‘adipokine’ be universally
reflect a protein found in the circulation, or (iii) if a
adopted to describe a protein that is secreted from (and
novel gene, the derived protein should contain a signal
sequence. When expression is first identified in adipose
Secretion is the critical characteristic of an adipokine,
tissue itself, it is essential to determine whether that
and we emphasise this since the term has also been used
expression occurs within mature adipocytes or in the
in connection with other adipocyte proteins such as adipo-
other cells that constitute the tissue, either histologically
nutrin (Wiesner et al. 2004), which is a transmembrane
(in situ hybridisation) or by separation of the adipocytes
protein rather than a secretory product. ‘Adipokine’ has
from the stromal vascular fraction by collagenase diges-
also been employed to describe a protein that is secreted
tion. Equally, expression in an adipocyte clonal line
by adipose tissue, rather than by adipocytes. However, it
needs to be verified for the native tissue. Gene expression
is preferable to restrict the term to those proteins that are
must, of course, be followed by detection of the encoded
released by adipocytes themselves. The principal reason
for this is that cells such as macrophages which also secrete
When a protein which is present in the circulation, or
protein signals are found in a number of organs, as well as
recognised to be secreted from other tissues, is synthesised
being present in adipose tissue (Weisberg et al. 2003; Xu
in adipocytes there is an a priori case for it being con-
et al. 2003). There is, therefore, a lack of specificity in
sidered an adipokine. Nevertheless, secretion from the adi-
giving the secreted proteins a special designation when
pocyte needs to be directly demonstrated before such a
such cells happen to be within fat depots, even though
protein should be accepted as a genuine adipokine.
their presence may well be of considerable functionalimportance.
There is also a question of whether the term adipokine
should be restricted to proteins released from white adipo-
The total number of adipokines, both documented and puta-
cytes, or include those secreted from brown adipocytes.
tive, is now well over fifty; the main functional categories
White and brown adipocytes are functionally different, of
are summarised in The earliest to be identified was
course, with the latter being specialised for the production
in practice the enzyme lipoprotein lipase, responsible for
of heat through the presence of the mitochondrial uncou-
the hydrolysis of circulating triacylglycerols to NEFA;
pling protein, UCP-1. In practice, no proteins appear to
this was followed in the mid 1980s by adipsin, a serine pro-
be secreted from brown adipocytes which are not also
tease and part of the alternative complement pathway
released by white fat cells, so in effect this may not be
(Cook et al. 1985, 1987). The key development which led
to the current focus on adipose tissue as a major site of
Given the current focus on ‘omic’ approaches in biology
the secretion of protein signals was, however, the discovery
(genome, transcriptome, proteome, metabolome/metabo-
in 1994 of leptin (Zhang et al. 1994). Leptin, which is a
nome), the totality of secreted proteins may be described
16 000 molecular-weight cytokine-like hormone with a
as constituting the adipokinome. Proteins are, however,
wide range of biological functions, established adipocytes
clearly not the only class of molecule secreted from adipo-
cytes. In addition to fatty acids, which quantitatively aremuch the largest secretory product, there are other lipidsubstances, including cholesterol, steroid hormones, prosta-glandins and prostanoids, and retinol (neither retinol norcholesterol are actually synthesised within adipocytes, butare stored and released). The lipid substances and adipoki-nome together can be said to constitute the ‘secretome’ ofthe adipocyte.
The identification of a protein as an adipokine requires thatsecretion from adipocytes be demonstrated. In practice, thiswill generally reflect selective release from adipocytes invitro. This may be either from freshly isolated adipocytes,or adipocytes derived by differentiation from fibroblastic
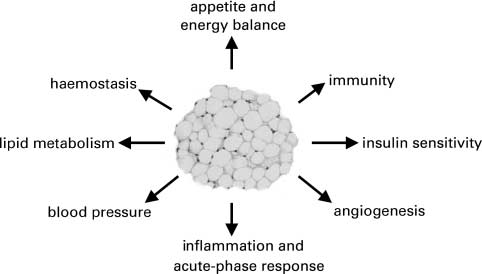
Fig. 1. Adipokines classified by functional role.
Role of white adipose tissue in inflammation
The diversity of the adipokines both in terms of
than adipose tissue, primarily the liver (and immune
protein structure and of putative function, is considerable
cells). The second explanation is that WAT is secreting
(Fru¨hbeck et al. 2001; Trayhurn & Beattie, 2001). The
factors that stimulate the production of inflammatory mar-
group includes: classical cytokines (e.g., TNFa, IL-6, IL-
kers from the liver and other organs; this may well be the
8), growth factors (e.g., transforming growth factor-b;
case with CRP, where it is argued that hepatic production
TGF-b) and proteins of the alternative complement
is stimulated by increased IL-6 from the expanded fat mass
system (e.g., adipsin, acylation-stimulating protein). The
of the obese (Yudkin et al. 2000; Yudkin, 2003). The third
group also includes: proteins involved in vascular haemo-
possibility is that adipocytes themselves are the immediate
stasis (e.g., plasminogen activator inhibitor-1 (PAI-1),
source of some, or most, of these inflammatory markers,
tissue factor), the regulation of blood pressure (angiotensi-
raised circulating levels in obesity reflecting production
nogen), lipid metabolism (e.g., retinol-binding protein,
from the increased WAT mass. There is also, of course,
cholesteryl ester transfer protein), glucose homeostasis
the possibility of there being a combination of production
(e.g., adiponectin, possibly resistin) and angiogenesis
(e.g., vascular endothelial growth factor; VEGF), as well
From the perspective of adipose tissue biology, a key
as acute-phase and stress responses (e.g., haptoglobin,
question is whether adipocytes (or adipose tissue) directly
contribute to the raised circulating levels of specific inflam-
From the wide range of protein signals and factors
matory markers and, if so, to what extent? Although
already identified, it is evident that WAT is a secretory
obtaining quantitative information on the contribution
and endocrine organ of considerable complexity which is
from particular cells within adipose tissue is difficult, the
highly integrated into the overall physiological and meta-
issue that can be readily addressed is whether adipocytes
bolic control systems of mammals. It is not easy to put for-
express certain inflammatory genes and their encoded pro-
ward a coherent framework for why such a diversity of
teins secreted. Recent reports demonstrating that WAT is
factors is secreted by white adipocytes. However, one
infiltrated by macrophages in obesity clearly suggest that
hypothesis would be that the various factors may relate
the non-adipocyte fraction may be a significant component
ultimately to the central lipid storage and release function
of the inflammatory state within adipose tissue (Weisberg
of the tissue (Trayhurn & Beattie, 2001). A corollary to
the secretion of such a wide range of adipokines is thatWAT has an extensive system for communication with
other tissues and organs. Co-culture studies have indicated,
Tumour necrosis factor-a and interleukin-6
for example, that adipocytes directly signal to other tissuessuch as skeletal muscle and the adrenal cortex (Dietze et al.
Several inflammatory cytokines are now recognised to be
2002; Ehrhart-Bornstein et al. 2003). There is also, in par-
expressed in, and secreted by, white adipocytes, the first
ticular, a distinct cross-talk between white adipocytes and
to be identified being TNFa (Hotamisligil et al. 1993).
the brain through leptin and the sympathetic nervous
TNFa expression in WAT was initially demonstrated in
system (Rayner & Trayhurn, 2001).
rodents, and found to be markedly increased in obesemodels (Hotamisligil et al. 1993). From this it was pro-posed that TNFa is linked to the development of insulin
resistance. The cytokine has been extensively examined
An important recent development in our understanding of
in relation to insulin action, and multiple effects have
obesity is the emergence of the concept that it (and dia-
been described, including the inhibition of the insulin
betes) is characterised by a state of chronic low-grade
receptor signalling pathway (Coppack, 2001; Hotamisligil,
inflammation (Yudkin et al. 1999; Das, 2001; Festa et al.
2003). In man, the secretion of TNFa is reported to be
2001; Engstro¨m et al. 2003). The basis for this view is
mainly due to the cells of the stromal vascular and
that increased circulating levels of several markers of
matrix fractions, including the macrophages, despite the
fact that previously most of the mRNA for TNFa was
acute-phase proteins, are elevated in the obese; these mar-
thought to be found within the adipocytes themselves
kers include IL-6, the TNFa system, C-reactive protein
(Weisberg et al. 2003; Fain et al. 2004a). An apparent
(CRP) and haptoglobin (Das, 2001; Bullo´ et al. 2003).
The implications in terms of the site of inflammation
human WAT is evident for several adipokines in recent
itself, whether systemic or local, are unclear. Nevertheless,
reports by Fain et al. (2004a,b) and requires further
it is increasingly evident that the inflammatory state may
be causal in the development of insulin resistance and
TNFa is a powerful local regulator within adipose
the other disorders associated with obesity, such as hyper-
tissue, acting in both an autocrine and a paracrine
lipidaemia and the metabolic syndrome (Hotamisligil,
manner to influence a range of processes, including apop-
2003; Yudkin, 2003). While the general assumption is
tosis (Prins et al. 1997; Coppack, 2001). There appears
that inflammation is consequent to obesity, it has been
to be a hierarchy of cytokines within WAT, with TNFa
suggested that obesity is in fact a result of inflammatory
playing a pivotal role in relation to the production of sev-
eral cytokines and other adipokines (Coppack, 2001). Thus,
A central question is the origin of the inflammatory mar-
for example, TNFa is a key regulator of the synthesis of
kers in obesity, and there are three possibilities. The first is
IL-6, of the acute-phase protein, haptoglobin (Chiellini
that it reflects production and release from organs other
et al. 2002; Oller do Nascimento et al. 2004), and of the
Fig. 2. Inflammatory and acute-phase response proteins secreted from adipocytes. The proteins in red have been clearly identified as adipo-kines (i.e. shown to be released by adipocytes) while those in blue are putative adipokines. CRP, C-reactive protein; IL, interleukin; NGF,nerve growth factor; PAI-1, plasminogen activator inhibitor-1; TGF-b, transforming growth factor-b; TNF-a, tumour necrosis factor-a; MCP-1,monocyte chemoattractant protein 1.
neurotrophin, nerve growth factor (NGF; Peeraully et al.
This cytokine stimulates the production of IL-6 and IL-8
2004). The extent to which TNFa produced in WAT is
from endothelial cells (Starnes et al. 2002). However, its
released into the circulation has been a matter of debate,
release from adipocytes has not been documented, so at pre-
but a relationship between the plasma TNFa system
sent it can only be considered as a putative adipokine.
(including the soluble receptors) and indices of obesityhas been reported (Bullo´ et al. 2003).
The other cytokine that has been the subject of major
interest in WAT is IL-6. It is expressed in, and secreted
There are a number of acute-phase proteins whose plasma
by, adipocytes and although it has local actions within
concentration increases substantially during the early
the tissue, it is released into the circulation (Mohamed-
stages of the inflammatory response, and a small number
Ali et al. 1997). Both plasma levels of IL-6 and expression
where the level falls (Gabay & Kushner, 1999). Several
in WAT are elevated in obesity and insulin resistance
of these proteins are now recognised as adipokines, with
(Mohamed-Ali et al. 1997; Bastard et al. 2000; Vozarova
adipose tissue being a potential contributor (either major
et al. 2001). It has been proposed that IL-6 has direct cen-
or minor) to the raised circulating levels in obesity;
tral actions, as IL-6 receptors are found in the hypothala-
mus in mice (Mohamed-Ali et al. 1997; Wallenius et al.
Expression of the genes encoding some acute-phase reac-
2002). As such, it is a candidate molecule for conveying
tants, such as a1-acid glycoprotein and 24p3, has been
information from adipocytes to the hypothalamus in the
observed in either adipocyte cell culture or in murine tis-
regulation of energy balance, additional to leptin.
sues (Soukas et al. 2000; Lin et al. 2001), but secretionas such has not yet been reported.
Although there has been considerable focus on TNFa and
IL-6, several other cytokines and related factors are syn-
PAI-1 is an important factor in the maintenance of vascular
thesised within adipose tissue, including IL-1b, TGF-b
haemostasis, inhibiting the activation of plasminogen, the
and, of course, leptin. Recent reports have included IL-8,
precursor of plasmin, which is involved in the breakdown
IL-10 and IL-17D. The IL-8 gene is expressed in human
of fibrin (Mutch et al. 2001). The expression and secretion
adipocytes and the protein released from both fat cells
of PAI-1 by adipocytes, both rodent and human, is well
and adipose tissue fragments (Bruun et al. 2000, 2001).
documented (Lundgren et al. 1996; Eriksson et al. 1998;
IL-1b and TNFa stimulate IL-8 release, while dexametha-
Cigolini et al. 1999; Mutch et al. 2001). The circulating
sone is inhibitory (Bruun et al. 2001). As with IL-6, the
level of PAI-1 is increased in obesity and synthesis in
plasma level of IL-8 is increased in obesity (Straczkowski
WAT is also raised (Alessi et al. 2000). This has led to
the view that adipose tissue is the major source of the ele-
Circulating levels of IL-18 are also increased in obesity
vated PAI-1 levels in the obese (Lundgren et al. 1996;
and fall with weight reduction (Esposito et al. 2002). As a
Samad et al. 1996; Alessi et al. 2000). As the risk of
consequence, it is speculated that WAT is a probable site
atherothrombotic disease is increased in obesity, this is a
of production of IL-18. Although there are no published
potent example of how the co-morbidities, such as diabetes
reports on this, we have recently observed IL-18 gene
and cardiovascular risk, associated with a high body fat can
expression in human WAT (IS Wood and P Trayhurn, unpub-
be directly linked to alterations in the production of
lished results). Similarly, the levels of IL-10, an anti-inflam-
specific adipokines. In addition to its role in haemostasis,
matory cytokine, are also raised in the obese (Esposito et al.
PAI-1 is also an acute-phase response protein, the levels
2003); the secretion of IL-10 from human adipocytes, as well
rising in inflammation (Gabay & Kushner, 1999).
as from the stromal vascular fraction and tissue matrix ofhuman fat depots, has been reported (Fain et al. 2004b).
The expression of IL-17D, believed to be the last member
of the IL-17 family to be identified, has recently been
Several studies have now reported that the haptoglobin
described in (human?) adipocytes (Starnes et al. 2002).
gene is expressed in murine adipose tissue (Friedrichs
Role of white adipose tissue in inflammation
et al. 1995; Chiellini et al. 2002; Oller do Nascimento et al.
(IS Wood and P Trayhurn, unpublished results). It should
2004). Similarly, gene expression has also been shown in
be noted that murine CRP is not regarded as an acute-
human WAT (Oller do Nascimento et al. 2004). A very
phase protein due to its very low expression (Volanakis,
recent study has demonstrated direct release of haptoglobin
from human adipose tissue explants (Fain et al. 2004a) and
Very low levels of expression would suggest that adipo-
release of haptoglobin into the medium has been observed
cytes are unlikely to be a significant direct contributor to
in 3T3-L1 adipocytes by a proteomic approach (Kratch-
circulating CRP levels. However, IL-6 is secreted by adi-
marova et al. 2002). Both transgenic studies and studies
pose tissue in increased amounts in obesity, as noted ear-
on 3T3-L1 adipocytes indicate that TNFa is a key factor
lier, and this is the major cytokine regulating the hepatic
in the stimulation of haptoglobin expression (Chiellini
production of CRP (Heinrich et al. 1990; Yudkin et al.
et al. 2002; Chinetti et al. 2003; Oller do Nascimento
2000). Thus WAT may be a major player in the raised cir-
et al. 2004), with IL-6 also being stimulatory.
culating levels of CRP in obesity, but through the indirect
Stimulation of the PPARg nuclear receptor through the
administration of the thiazolidienedione, rosiglitazone,strongly inhibits haptoglobin gene expression (Oller do
Nascimento et al. 2004). This is consistent with the emer-ging view that PPARs have substantial anti-inflammatory
There are several other adipokines involved in the inflam-
actions (Moller & Berger, 2003). Indeed, several other
matory response that are neither cytokines nor acute-phase
inflammation-related adipokines are down regulated by
proteins. Interestingly, adiponectin (also known as Acrp30,
PPARg ligands, including TNFa, leptin and NGF, while
AdipoQ, ApM1 or GBP28), which is synthesised only in
there is evidence that adiponectin is up regulated (Moller
adipose tissue, appears to have an anti-inflammatory
effect, inhibiting phagocytic activity and TNFa productionin macrophages (Ouchi et al. 1999; Yokota et al. 2000). This adipokine is now very much centre stage; this is
partly because, in contrast to many other adipokines, its
Serum amyloid A (SAA), the precursor to amyloid A pro-
expression and release fall in obesity (Arita et al. 1999;
tein found in secondary amyloid plaques, consists of a
Hotta et al. 2000). However, the principal reason for the
family of apolipoproteins which bind to, and substitute
recent focus on adiponectin is the evidence that it is
for, apo A-I in HDL. These apolipoproteins are expressed
involved in modulating insulin sensitivity (Berg et al.
as either major acute-phase reactants SAA, or constitutive
2001; Yamauchi et al. 2001), as well as having anti-athero-
SAA, the functions of which are largely unknown. How-
geneic properties (Engeli et al. 2003).
ever, a few clinically important functions have been
We have recently observed that the target-derived neuro-
suggested that include pro-inflammatory and anti-inflam-
trophin, NGF, is synthesised by the main adipose tissue
matory roles. A number of genes have now been identified
depots in both rodents and man, and is secreted from
in man and mice which share very similar sequence iden-
white adipocytes (Peeraully et al. 2004). This protein,
tities and genomic organisation (Uhlar & Whitehead,
which was the first of the family of neurotrophins to be dis-
1999). These genes are up regulated by pro-inflammatory
covered, was originally linked to the growth and survival
cytokines such as TNFa and IL-6, as well as by glucocor-
of sympathetic neurones; however, although not a cyto-
ticoids. The expression of SAA, as with other acute-phase
kine, it is now recognised as also being involved in
reactants, is predominantly in the liver. However, extrahe-
immune and inflammatory responses (Levi-Montalcini
patic expression of SAA, including adipocytes, has been
et al. 1996; Vega et al. 2003). Indeed, NGF is expressed
reported; expression and release of SAA3 occurs in
in adipocytes specifically associated with wound healing
murine adipocytes, and this is up regulated under hypergly-
and with atherosclerotic lesions (Hasan et al. 2000; Chal-
caemic conditions (Lin et al. 2001).
dakov et al. 2001). Importantly, TNFa has a strong stimu-latory effect on NGF gene expression and NGF releasefrom 3T3-L1 adipocytes, suggesting that the neurotrophin
is an inflammatory response protein in adipose tissue (Peer-
The circulating level of CRP rises with BMI (Visser et al.
1999; Pannacciulli et al. 2001; Bullo´ et al. 2003), and elev-ated levels of this inflammatory marker have been associ-
Why inflammation in obesity: a response to hypoxia?
ated with both obesity and diabetes, falling with weightloss (Tchernof et al. 2002). There is evidence from a
Much attention has been directed towards unravelling the
study using real-time PCR that the gene encoding CRP is
pathological and clinical implications of inflammation in
expressed in adipose tissue, an inverse correlation between
obesity, and establishing the links with insulin resistance
the levels of the mRNA for CRP and adiponectin being
and other metabolic disorders – the metabolic syndrome.
apparent (Ouchi et al. 2003). This raises the possibility
However, a central issue is why does WAT release pro-
that adipose tissue contributes directly to the circulating
inflammatory cytokines and acute-phase proteins, and why
pool of CRP. Unfortunately, it is not clear whether CRP
do these rise sharply as fat mass increases? Linked to this
expression in adipocytes is in practice significant; our
is the question of why obesity should be associated
own studies using conventional RT-PCR have found that
with chronic low-grade inflammation. Adipose tissue may,
there is little expression in human WAT, or adipocytes
of course, be contributing inflammation-related factors
to a specific site of inflammation in an organ (or organs)
occurs in both the adipocytes and in the stromal vascular
elsewhere, or as part of a systemic state of inflammation.
cells, and in the WAT of obese (ob/ob) mice the level of
However, an alternative view is that the inflammatory
the mRNA is markedly increased compared with lean sib-
state is mainly within WAT itself. If the inflammatory
lings (L Hunter, IS Wood and P Trayhurn, unpublished
response is primarily local to adipose tissue, at least in
results). Thus a link between the increased WAT mass in
terms of its initiation, then the elevated circulating levels
obesity, adipocyte hypoxia, inflammation and the stimu-
of inflammation-related products may in effect reflect spil-
lation of angiogenesis is plausible.
lover from the tissue, and the link with insulin resistancewould be an incidental consequence.
In the absence of any specific indication to the contrary,
the parsimonious view is that the secretion of inflammatory
We have focused in the present paper on the role of WAT
cytokines and acute-phase proteins by adipocytes in obesity
in inflammatory responses. There have, however, been a
relates to events within WAT itself. If this is correct, what
number of other important recent developments in the adi-
could be the rationale for a local effect? A possible expla-
pokine field. For example, a recent report has indicated that
nation is that it is a response to hypoxia in areas of the fat
human white adipocytes secrete mineralocorticoid-releas-
depots as the tissue mass increases during the progressive
ing factors, aldosterone secretion by adrenocortical cells
development of obesity. The sequence of events might be
being stimulated (Ehrhart-Bornstein et al. 2003). These
that as the tissue expands, the vasculature (which is less
results indicate a direct link between obesity and hyperten-
extensive in WAT than in brown fat) is insufficient to main-
sion through adipose tissue regulating mineralocorticoid
tain normoxia throughout the organ. Clusters of adipocytes
then become relatively hypoxic, and an inflammatory
There is continuing interest in whether adipocytes
response ensues which serves to increase blood flow and
secrete centrally acting signals in the regulation of appetite
to stimulate angiogenesis. This has some parallels with
and energy balance additional to leptin, as noted earlier. An
tumour growth in cancer. That vascular development is an
initial candidate for such a role was fasting-induced adi-
important issue in WAT function is indicated by the fact
pose factor (also known as PPARg angiopoietin-related
that adipose tissue mass is sensitive to angiogenesis inhibi-
gene) (Kersten et al. 2000; Yoon et al. 2000). Fasting-
tors and can be regulated by its vasculature (Rupnick et al.
induced adipose factor is an angiopoietin-related protein,
2002), and that several angiogenic factors are secreted by
the expression of which is strongly stimulated by fasting
adipocytes (Claffey et al. 1992; Rupnick et al. 2002; Lol-
in a manner which may be reciprocal to leptin. Fasting-
mede et al. 2003). These factors include recognised angio-
induced adipose factor is potentially of considerable inter-
genic signals (VEGF, PAI-1 and leptin), as well as
est, not only as a putative signal in energy balance, but as
putative signals such as metallothionein and haptoglobin.
part of the adaptive response to nutritional deprivation. In
A pivotal signal in the cellular response to hypoxia is
this regard, since it is induced by fasting, it is a positive
signal of the fasted state, in contrast to leptin which signals
factor is a heterodimeric protein consisting of a and b sub-
food deprivation through a reduction in its expression
units. The b subunit is constitutively expressed, but the a
subunit is highly induced by hypoxia leading to the formation
An intriguing report using DNA microarrays for gene
of functional HIF-1 (Semenza, 2001; Wenger, 2002; Ho¨pfl
expression profiling of human visceral adipose tissue has
et al. 2004). Hypoxia is characteristic of tumours, where
suggested that a constellation of neuroendocrine factors
HIF-1 expression is increased, as well as in other disorders
may be produced by white adipocytes (Yang et al. 2003).
such as ischaemic heart disease (Binley et al. 2003; Ho¨pfl
These factors included cholecystokinin, neurotensin and
et al. 2004). A number of genes are regulated by HIF-1,
neuropeptide Y. If correct, it suggests that the adipocyte
which acts as a central controller of oxygen-regulated gene
is even more remarkable as a secretory cell than currently
expression. The transcription factor is stabilised and its
envisaged; indeed, it would be a veritable powerhouse in
expression stimulated by cytokines such as TNFa and IL-
the secretion of neuroendocrine signals. However, our
1b (Hellwig-Bu¨rgel et al. 1999). The target genes for HIF-
attempts to identify transcripts of some of these genes
1 include VEGF and PAI-1 (Ho¨pfl et al. 2004). In addition,
using conventional RT – PCR have proved unsuccessful
there is now evidence for the transcriptional activation of
(L Hunter, IS Wood and P Trayhurn, unpublished results),
leptin through HIF-1a in response to hypoxia (Ambrosini
and the reported expression may in practice reflect the pro-
blems inherent in setting a suitable (arbitrary) threshold in
Immunoreactive HIF-1a has been reported in 3T3-
microarray studies when comparisons between groups are
F442A adipocytes and hypoxia results in an increase in
not part of the experimental paradigm.
the amount of the protein in the cultured cells (Lolmedeet al. 2003). Furthermore, hypoxia leads to an induction
of leptin and VEGF expression in these adipocytes, raisingthe likelihood that a low oxygen tension leads to the stimu-
The biology of WAT seems ever more rich and complex.
lation of angiogenesis in adipose tissue through the HIF-1
The family of adipokines is increasing rapidly, these
proteins being highly diverse in structure and in function.
HIF-1a expression is not just a feature of cultured adipo-
A critical issue is the physiological role that many of
cytes, since we have recently observed that the HIF-1a
them play. The role may be local (through an autocrine
gene is expressed in mouse WAT depots. Expression
or paracrine function) or endocrine, or involve multiple
Role of white adipose tissue in inflammation
levels (as with leptin and IL-6). An important development
Bullo´ M, Garcia-Lorda P, Megias I & Salas-Salvado J (2003) Sys-
is the recognition of the extensive and direct involvement
temic inflammation, adipose tissue tumor necrosis factor, and
of white adipocytes in inflammation and the acute-phase
leptin expression. Obesity Res 11, 525 – 531.
response. In some cases it seems probable that adipocytes
Chaldakov GN, Stankulov IS, Fiore M, Ghenev PI & Aloe L
(2001) Nerve growth factor levels and mast cell distribution
contribute substantially to the raised circulating levels of
in human coronary atherosclerosis. Atherosclerosis 159,
particular pro-inflammatory cytokines and acute-phase
reactants in obesity, such as IL-6, PAI-1 and haptoglobin.
Chiellini C, Bertacca A, Novelli SE, et al. (2002) Obesity modu-
In others, adipose tissue appears to have an indirect role;
lates the expression of haptoglobin in the white adipose tissue
for example, hepatic production of CRP may be stimulated
via TNFa. J Cell Physiol 190, 251 – 258.
by the increased release of IL-6 from adipocytes.
Chinetti G, Fruchart JC & Staels B (2003) Peroxisome prolifera-
The inflammatory state that is characteristic of obesity
tor-activated receptors and inflammation: from basic science to
may primarily relate to local events within adipose
clinical applications. Int J Obesity 27, Suppl. 3, S41 – S45.
tissue, raised circulating levels of inflammatory cytokines
Cigolini M, Tonoli M, Borgato L, et al. (1999) Expression of
and acute-phase proteins that are adipokines reflecting
plasminogen activator inhibitor-1 in human adipose tissue: arole for TNF-a? Atherosclerosis 143, 81 – 90.
spillover from the tissue. Hypoxia consequent upon the
Claffey KP, Wilkison WO & Spiegelman BM (1992) Vascular
expansion of white fat mass in advance of angiogenesis
endothelial growth factor. Regulation by cell differentiation
could be a key trigger for the inflammation-related events
and activated second messenger pathways. J Biol Chem 267,
cytokines may lead to a direct stimulation of angiogenic
Cook KS, Groves DL, Min HY & Spiegelman BM (1985) A
factors, such as VEGF and leptin, as well as through the
activation of HIF-1, the central controller of the cellular
encodes a novel serine protease homologue. Proc Natl Acad
Cook KS, Min HY, Johnson D, Chaplinsky RJ, Flier JS, Hunt CR
& Spiegelman BM (1987) Adipsin: a circulating serine pro-
tease homolog secreted by adipose tissue and sciatic nerve. Science 237, 402 – 405.
We thank Dr Mo`nica Bullo´ Bonet for her helpful com-
Coppack SW (2001) Pro-inflammatory cytokines and adipose
ments on the manuscript. Our work is funded by grants
tissue. Proc Nutr Soc 60, 349 – 356.
from the BBSRC, European Union and the University of
Das UN (2001) Is obesity an inflammatory condition? Nutrition
Dietze D, Koenen M, Rohrig K, Horikoshi H, Hauner H & Eckel
J (2002) Impairment of insulin signaling in human skeletalmuscle cells by co-culture with human adipocytes. Diabetes
Ahima RS, Prabakaran D, Mantzoros C, Qu DQ, Lowell B,
Ehrhart-Bornstein M, Lamounier-Zepter V, Schraven A, Langen-
Maratos-Flier E & Flier JS (1996) Role of leptin in the neuro-
bach J, Willenberg HS, Barthel A, Hauner H, McCann SM,
endocrine response to fasting. Nature 382, 250 – 252.
Scherbaum WA & Bornstein SR (2003) Human adipocytes
Alessi MC, Bastelica D, Morange P, Berthet B, Leduc I, Verdier
secrete mineralocorticoid-releasing factors. Proc Natl Acad
M, Geel O & Juhan-Vague I (2000) Plasminogen activator
inhibitor 1, TGFb1, and BMI are closely associated in human
Engeli S, Feldpausch M, Gorzelniak K, Hartwig F, Heintze U,
adipose tissue during morbid obesity. Diabetes 49, 1374 – 1380.
Janke J, Mohlig M, Pfeiffer AFH, Luft FC & Sharma AM
Ambrosini G, Nath AK, Sierra-Honigmann MR & Flores-Riveros
(2003) Association between adiponectin and mediators of
J (2002) Transcriptional activation of the human leptin gene in
inflammation in obese women. Diabetes 52, 942 – 947.
response to hypoxia. Involvement of hypoxia-inducible factor
Engstro¨m G, Hedblad B, Stavenow L, Lind P, Janzon L &
1. J Biol Chem 277, 34601 – 34609.
Lindga¨rde F (2003) Inflammation-sensitive plasma proteins
Arita Y, Kihara S, Ouchi N, et al. (1999) Paradoxical decrease of
an adipose-specific protein, adiponectin, in obesity. Biochem
Eriksson P, Reynisdottir S, Lonnqvist F, Stemme V, Hamsten A
Bastard JP, Jardel C, Bruckert E, Blondy P, Capeau J, Laville M,
& Arner P (1998) Adipose tissue secretion of plasminogen
Vidal H & Hainque B (2000) Elevated levels of interleukin 6
activator inhibitor-1 in non-obese and obese individuals.
are reduced in serum and subcutaneous adipose tissue of
obese women after weight loss. J Clin Endocrinol Metab 85,
Esposito K, Pontillo A, Ciotola M, Di Palo C, Grella E, Nicoletti
G & Giugliano D (2002) Weight loss reduces interleukin-18
Berg AH, Combs TP, Du X, Brownlee M & Scherer PE
levels in obese women. J Clin Endocrinol Metab 87,
(2001) The adipocyte-secreted protein Acrp30 enhances
hepatic insulin action. Nature Med 7, 947 – 953.
Esposito K, Pontillo A, Giugliano F, Giugliano G, Marfella R,
Binley K, Kan O, White J & Naylor S (2003) Exploiting the
Nicoletti G & Giugliano D (2003) Association of low interleu-
hypoxia response. Curr Opin Mol Ther 5, 650 – 656.
kin-10 levels with the metabolic syndrome in obese women.
Bruun JM, Pedersen SB & Richelsen B (2000) Interleukin-8
J Clin Endocrinol Metab 88, 1055 – 1058.
production in human adipose tissue. Inhibitory effects of
Fain JN, Bahouth SW & Madan AK (2004a) Haptoglobin release
anti-diabetic compounds, the thiazolidinedione ciglitazone
by human adipose tissue in primary culture. J Lipid Res 45,
and the biguanide metformin. Horm Metab Res 32, 537 – 541.
Bruun JM, Pedersen SB & Richelsen B (2001) Regulation of
Fain JN, Madan AK, Hiler ML, Cheema P & Bahouth SW
interleukin 8 production and gene expression in human adipose
(2004b) Comparison of the release of adipokines by adipose
tissue in vitro. J Clin Endocrinol Metab 86, 1267 – 1273.
tissue, adipose tissue matrix, and adipocytes from visceral
and subcutaneous abdominal adipose tissues of obese humans.
Mohamed-Ali V, Goodrick S, Rawesh A, Katz DR, Miles JM,
Yudkin JS, Klein S & Coppack SW (1997) Subcutaneous
Festa A, D’Agostino R Jr, Williams K, Karter AJ, Mayer-Davis
adipose tissue releases interleukin-6, but not tumor necrosis
EJ, Tracy RP & Haffner SM (2001) The relation of body fat
factor-alpha, in vivo. J Clin Endocrinol Metab 82, 4196 – 4200.
mass and distribution to markers of chronic inflammation. Int
Mohamed-Ali V, Pinkney JH & Coppack SW (1998) Adipose
tissue as an endocrine and paracrine organ. Int J Obesity 22,
Friedrichs WE, Navarijo-Ashbaugh AL, Bowman BH & Yang F
(1995) Expression and inflammatory regulation of haptoglobin
Moller DE & Berger JP (2003) Role of PPARs in the regulation
gene in adipocytes. Biochem Biophys Res Commun 209,
of obesity-related insulin sensitivity and inflammation. Int
J Obesity 27, Suppl. 3, S17 – S21.
Fru¨hbeck G, Go´mez-Ambrosi J, Muruzabal FJ & Burrell MA
Mutch NJ, Wilson HM & Booth NA (2001) Plasminogen activa-
(2001) The adipocyte: a model for integration of endocrine
tor inhibitor-1 and haemostasis in obesity. Proc Nutr Soc 60,
and metabolic signaling in energy metabolism regulation. Am
Oller do Nascimento C, Hunter L & Trayhurn P (2004) Regu-
Funahashi T, Nakamura T, Shimomura I, Maeda K, Kuriyama H,
lation of haptoglobin gene expression in 3T3-L1 adipocytes
Takahashi M, Arita Y, Kihara S & Matsuzawa Y (1999) Role
by cytokines, catecholamines, and PPARg. Biochem Biophys
of adipocytokines on the pathogenesis of atherosclerosis in
visceral obesity. Intern Med 38, 202 – 206.
Ouchi N, Kihara S, Arita Y, et al. (1999) Novel modulator for
Gabay C & Kushner I (1999) Acute-phase proteins and other
endothelial adhesion molecules - adipocyte-derived plasma
systemic responses to inflammation. New Engl J Med 340,
protein adiponectin. Circulation 100, 2473 – 2476.
Ouchi N, Kihara S, Funahashi T, et al. (2003) Reciprocal associ-
Hasan W, Zhang R, Liu M, Warn JD & Smith PG (2000)
ation of C-reactive protein with adiponectin in blood stream
Coordinate expression of NGF and alpha-smooth muscle
and adipose tissue. Circulation 107, 671 – 674.
actin mRNA and protein in cutaneous wound tissue of
Pannacciulli N, Cantatore FP, Minenna A, Bellacicco M,
developing and adult rats. Cell Tissue Res 300, 97 – 109.
Giorgino R & De Pergola G (2001) C-reactive protein is
Heinrich PC, Castell JV & Andus T (1990) Interleukin-6 and the
independently associated with total body fat, central fat, and
acute phase response. Biochem J 265, 621 – 636.
insulin resistance in adult women. Int J Obesity 25,
Hellwig-Bu¨rgel T, Rutkowski K, Metzen E, Fandrey J &
Jelkmann W (1999) Interleukin-1b and tumor necrosis factor-
Peeraully MR, Jenkins JR & Trayhurn P (2004) NGF gene
a stimulate DNA binding of hypoxia-inducible factor-1.
expression and secretion in white adipose tissue: regulation in
3T3-L1 adipocytes by hormones and inflammatory cytokines.
Ho¨pfl G, Ogunshola O & Gassmann M (2004) HIFs and tumors -
causes and consequences. Am J Physiol 286, R608 – R623.
Prins JB, Niesler CU, Winterford CM, Bright NA, Siddle K,
Hotamisligil GS (2003) Inflammatory pathways and insulin
Orahilly S, Walker NI & Cameron DP (1997) Tumor necrosis
action. Int J Obesity 27, Suppl. 3, S53 – S55.
factor-alpha induces apoptosis of human adipose cells.
Hotamisligil GS, Shargill NS & Spiegelman BM (1993) Adipose
expression of tumor necrosis factor-alpha - direct role in obes-
Rajala MW & Scherer PE (2003) Minireview: the adipocyte - at
ity-linked insulin resistance. Science 259, 87 – 91.
the crossroads of energy homeostasis, inflammation, and
Hotta K, Funahashi T, Arita Y, et al. (2000) Plasma concen-
atherosclerosis. Endocrinology 144, 3765 – 3773.
trations of a novel, adipose-specific protein, adiponectin, in
Rayner DV & Trayhurn P (2001) Regulation of leptin production:
type 2 diabetic patients. Arterioscl Thromb Vasc Biol 20,
sympathetic nervous system interactions. J Mol Med 79, 8 – 20.
Rupnick MA, Panigrahy D, Zhang C-Y, Dallabrida SM, Lowell
Kersten S, Mandard S, Tan NS, Escher P, Metzger D, Chambon
BB, Langer R & Folkman MJ (2002) Adipose tissue mass
P, Gonzalez FJ, Desvergne B & Wahli W (2000) Characteriz-
can be regulated through the vasculature. Proc Natl Acad Sci
ation of the fasting-induced adipose factor FIAF, a novel
peroxisome proliferator-activated receptor target gene. J Biol
Samad F, Yamamoto K & Loskutoff DJ (1996) Distribution and
regulation of plasminogen activator inhibitor-1 in murine adi-
Klaus S (2004) Adipose tissue as a regulator of energy balance.
pose tissue in vivo - induction by tumor necrosis factor-a and
lipopolysaccharide. J Clin Invest 97, 37 – 46.
Kratchmarova I, Kalume DE, Blagoev B, et al. (2002) A proteo-
Semenza GL (2001) HIF-1 and mechanisms of hypoxia sensing.
mic approach for identification of secreted proteins during the
Curr Opin Cell Biol 13, 167 – 171.
differentiation of 3T3-L1 preadipocytes to adipocytes. Mol
Soukas A, Cohen P, Socci ND & Friedman JM (2000) Leptin-
specific patterns of gene expression in white adipose tissue.
Levi-Montalcini R, Skaper SD, Dal Toso R, Petrelli L & Leon A
(1996) Nerve growth factor: from neurotrophin to neurokine.
Starnes T, Broxmeyer HE, Robertson MJ & Hromas R (2002)
Cutting edge: IL-17D, a novel member of the IL-17 family,
Lin Y, Rajala MW, Berger JP, Moller DE, Barzilai N & Scherer
stimulates cytokine production and inhibits hemopoiesis.
PE (2001) Hyperglycemia-induced production of acute phase
reactants in adipose tissue. J Biol Chem 276, 42077 – 42083.
Straczkowski M, Dzienis-Straczkowska S, Stepien A, Kowalska I,
Lolmede K, Durand de Saint Front V, Galitzky J, Lafontan M &
Szelachowska M & Kinalska I (2002) Plasma interleukin-8
Bouloumie A (2003) Effects of hypoxia on the expression of
concentrations are increased in obese subjects and related to
proangiogenic factors in differentiated 3T3-F442A adipocytes.
fat mass and tumor necrosis factor-a system. J Clin Endocrinol
Lundgren CH, Brown SL, Nordt TK, Sobel BE & Fujii S (1996)
Tchernof A, Nolan A, Sites CK, Ades PA & Poehlman ET (2002)
Elaboration of type-1 plasminogen activator inhibitor from
Weight loss reduces C-reactive protein levels in obese postme-
adipocytes - a potential pathogenetic link between obesity
nopausal women. Circulation 105, 564 – 569.
and cardiovascular disease. Circulation 93, 106 – 110.
Trayhurn P & Beattie JH (2001) Physiological role of adipose
Role of white adipose tissue in inflammation
tissue: white adipose tissue as an endocrine and secretory
fat plays a crucial role in the development of obesity-related
organ. Proc Nutr Soc 60, 329 – 339.
insulin resistance. J Clin Invest 112, 1821 – 1830.
Uhlar CM & Whitehead AS (1999) Serum amyloid A, the
Yamauchi T, Kamon J, Waki H, et al. (2001) The fat-derived hor-
major vertebrate acute-phase reactant. Eur J Biochem 265,
mone adiponectin reverses insulin resistance associated with
both lipoatrophy and obesity. Nature Med 7, 941 – 946.
Vega JA, Garcia-Suarez O, Hannestad J, Perez-Perez M &
Yang YS, Song HD, Li RY, Zhou LB, Zhu ZD, Hu RM, Han ZG
Germana A (2003) Neurotrophins and the immune system.
& Chen JL (2003) The gene expression profiling of human
visceral adipose tissue and its secretory functions. Biochem
Visser M, Bouter LM, McQuillan GM, Wener MH & Harris TB
Biophys Res Commun 300, 839 – 846.
(1999) Elevated C-reactive protein levels in overweight and
Yokota T, Oritani K, Takahashi I, et al. (2000) Adiponectin, a
obese adults. J Am Med Assoc 282, 2131 – 2135.
new member of the family of soluble defense collagens, nega-
Volanakis JE (2001) Human C-reactive protein: expression,
tively regulates the growth of myelomonocytic progenitors and
structure, and function. Mol Immunol 38, 189 – 197.
the functions of macrophages. Blood 96, 1723 – 1732.
Vozarova B, Weyer C, Hanson K, Tataranni PA, Bogardus C &
Yoon JC, Chickering TW, Rosen ED, Dussault B, Qin Y,
Pratley RE (2001) Circulating interleukin-6 in relation to
Soukas A, Friedman JM, Holmes WE & Spiegelman BM
adiposity, insulin action, and insulin secretion. Obesity Res 9,
(2000) Peroxisome proliferator-activated receptor gamma
target gene encoding a novel angiopoietin-related protein
Wallenius K, Wallenius V, Sunter D, Dickson SL & Jansson JO
associated with adipose differentiation. Mol Cell Biol 20,
decreases body fat in rats. Biochem Biophys Res Commun
Yudkin JS (2003) Adipose tissue, insulin action and vascular
disease: inflammatory signals. Int J Obesity 27, Suppl. 3,
Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL
& Ferrante AW Jr (2003) Obesity is associated with macro-
Yudkin JS, Kumari M, Humphries SE & Mohamed-Ali V (2000)
phage accumulation in adipose tissue. J Clin Invest 112,
Inflammation, obesity, stress and coronary heart disease: is
interleukin-6 the link? Atherosclerosis 148, 209 – 214.
Wenger RH (2002) Cellular adaptation to hypoxia: O2-sensing
Yudkin JS, Stehouwer CD, Emeis JJ & Coppack SW (1999)
protein hydroxylases, hypoxia-inducible transcription factors,
C-reactive protein in healthy subjects: associations with obes-
and O2-regulated gene expression. FASEB J 16, 1151– 1162.
ity, insulin resistance, and endothelial dysfunction: a potential
Wiesner G, Morash BA, Ur E & Wilkinson M (2004)
role for cytokines originating from adipose tissue? Arterioscler
pituitary gland but not in hypothalamus. J Endocrinol 180,
Zhang YY, Proenca R, Maffei M, Barone M, Leopold L &
Friedman JM (1994) Positional cloning of the mouse obese
Xu H, Barnes GT, Yang Q, et al. (2003) Chronic inflammation in
gene and its human homolog. Nature 372, 425 – 432.
LEA DETENIDAMENTE ESTE DOCUMENTO Y TENGA EN CUENTA LAS ORIENTACIONES ANTES DE ACUDIR A LA CONSULTA PRE-OPERATORIA DEL HOSPITAL DE OJOS DEBE MOSTRARLE A SU MÉDICO TRATANTE ESTE DOCUMENTO Y SI TIENE ALGUNA DUDA SE PUEDE COMUNICAR CON LA COORDINACIÓN QUE LE CORRESPONDA. ASPECTOS GENERALES: Es importante que sepa que usted no acude a operarse, sino a una evaluación pre-opera
GENEESMIDDELEN EN RIJVAARDIGHEID R.A. Bredewoud Hoofd medische zaken CBR Postbus 3014, 2280 GA Rijswijk Inleiding Het Centraal Bureau Rijvaardigheidsbewijzen (CBR) is als organisatie vooral bekend vanwege het toetsen van de rijvaardigheid: het rijexamen. Daarnaast toetst de afdeling Medische Zaken sinds 1951 echter ook de rijgeschiktheid: de lichamelijke en geestelijke geschiktheid
 accord them a collective name. The term initially intro-
pre-adipocytes – whether primary culture or from clonal
duced was ‘adipocytokines’ (Funahashi et al. 1999), and
cell lines. Alternative approaches, which are more challen-
this has been used extensively. Although the name has
ging, include detection of the protein in the venous drai-
merit, it is potentially misleading since there is an infer-
nage from WAT at a concentration higher than in arterial
ence that the adipocyte-secreted proteins are cytokines, or
cytokine-like. While this is the case for some, such as
The initial stage in the identification of a candidate adi-
TNFa and IL-6, it is clearly not so with the majority.
accord them a collective name. The term initially intro-
pre-adipocytes – whether primary culture or from clonal
duced was ‘adipocytokines’ (Funahashi et al. 1999), and
cell lines. Alternative approaches, which are more challen-
this has been used extensively. Although the name has
ging, include detection of the protein in the venous drai-
merit, it is potentially misleading since there is an infer-
nage from WAT at a concentration higher than in arterial
ence that the adipocyte-secreted proteins are cytokines, or
cytokine-like. While this is the case for some, such as
The initial stage in the identification of a candidate adi-
TNFa and IL-6, it is clearly not so with the majority.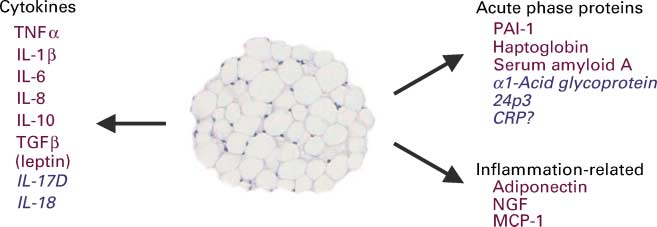 Fig. 2. Inflammatory and acute-phase response proteins secreted from adipocytes. The proteins in red have been clearly identified as adipo-kines (i.e. shown to be released by adipocytes) while those in blue are putative adipokines. CRP, C-reactive protein; IL, interleukin; NGF,nerve growth factor; PAI-1, plasminogen activator inhibitor-1; TGF-b, transforming growth factor-b; TNF-a, tumour necrosis factor-a; MCP-1,monocyte chemoattractant protein 1.
Fig. 2. Inflammatory and acute-phase response proteins secreted from adipocytes. The proteins in red have been clearly identified as adipo-kines (i.e. shown to be released by adipocytes) while those in blue are putative adipokines. CRP, C-reactive protein; IL, interleukin; NGF,nerve growth factor; PAI-1, plasminogen activator inhibitor-1; TGF-b, transforming growth factor-b; TNF-a, tumour necrosis factor-a; MCP-1,monocyte chemoattractant protein 1.