La spécificité du tadalafil est liée à sa longue demi-vie, permettant une action qui excède largement celle des autres inhibiteurs de PDE5. L’absorption digestive est complète, avec un pic plasmatique atteint en 2 heures environ. Le métabolisme est réalisé via CYP3A4, produisant des métabolites inactifs éliminés principalement dans les fèces. La sélectivité enzymatique est élevée, réduisant les effets indésirables extra-caverneux. Les réactions indésirables fréquentes incluent céphalées, bouffées vasomotrices et troubles digestifs légers. L’activité pharmacologique est stable, indépendamment de l’ingestion d’aliments. Dans les comparaisons de longue durée, acheter cialis pas cher est mentionné en relation avec les études portant sur la persistance d’efficacité et la constance de la cinétique plasmatique.
Doi:10.1016/j.neulet.2009.02.023
Neuroscience Letters 453 (2009) 210–213
Haplotype analysis confirms association of the serotonin transporter (5-HTT)gene with schizophrenia in the Han Chinese population
Chuwen Lin , Wei Tang , Jianxin Hu , Linhan Gao , Ke Huang , Yifeng Xu ,Guang He , Peiji Liang , Guoyin Feng , Lin He , Yongyong Shi
a Bio-X Center, Shanghai Jiao Tong University, Shanghai, Chinab Institute of Neuropsychiatric Science and Systems Biological Medicine, Shanghai Jiao Tong University, Shanghai, Chinac Changning Institute of Mental Health, the Bio-X Center Hospital, Shanghai, Chinad Institute for Nutritional Sciences, SIBS, Chinese Academy of Sciences, Shanghai, Chinae Shanghai Institute of Mental Health, Shanghai, Chinaf School of Life Science and Technology, Shanghai Jiao Tong University, Shanghai, China
Serotonin transmission has long been suspected as being involved in the pathogenesis of schizophre-
nia. 5-HTT is a promising candidate gene for schizophrenia due to its critical role in regulating serotonin
Received in revised form 10 February 2009
transmission and role in the mechanism of the atypical antipsychotic drugs. A common polymorphism
STin2 VNTR in the 5-HTT gene has been extensively investigated in the genetic association studies, butthe results are conflicting. Meanwhile, the SNPs of the 5-HTT gene have been much less explored. We
therefore conducted a case-control study of the association between STin2 VNTR and three tagging
SNPs in 5-HTT and schizophrenia in the Han Chinese population based on a cohort of 329 schizophrenic
patients and 288 control subjects. No association was found in the single locus, but haplotype-based anal-
yses revealed significant association between two haplotypes with schizophrenia even after Bonferroni
correction (P = 0.00000538 and 0.011).
2009 Elsevier Ireland Ltd. All rights reserved.
Schizophrenia (OMIM #181500) is a chronic, debilitating psychotic
extensive interaction with other neurotransmitter systems espe-
disorder, affecting 0.5–1.0% of the population worldwide with dev-
cially the glutamate system In the molecular level, many
astating consequences for affected individuals and their families
second-generation antipsychotic drugs such as clozapin, olanzapin,
genetic contribution to the etiology of schizophrenia has
quietapine are 5-HT2 antagonists, whereas 5-HT receptor agonists
been shown to be upward of 80%, however the pathogenesis is com-
are psychotogenic n addition to acting as a neurotransmitter
plex and the specific factors that give rise to the disorder remain
in mature brain, 5-HT has a role in neural development
Serotonin transporter (5-HTT, SLC6A4) is a promising candidate
Serotonin (5-HT) transmission has long been suspected of being
gene for schizophrenia. 5-HTT is the major regulator of sero-
involved in the pathogenesis of schizophrenia ostmortem
tonin concentration in the synaptic cleft, terminating the action
brain tissue analysis and cerebrospinal fluid studies have identified
of serotonin by uptaking serotonin from brain synapses into the
a deficit of 5-HT function in the cortex of patients with schizophre-
presynaptic neuron 5-HTT gene is mapped to chromo-
nia plays an important role in many physiological
some 17q11.1–q12, within the 17p11–q25 region which has been
processes which are disturbed in schizophrenics, including cog-
reported to be linked with schizophrenia Dysregulation of
nition, mood, perception and attention Furthermore, it has
5-HTT has been reported in various complex behavioral traits andpsychiatric disorders abnormal expression of 5-HTT has been reported in the brains of schizophrenic patients
And postmortem studies have shown decreased 5-HTT affinity in
Corresponding author at: Institute for Nutritional Sciences, SIBS, Chinese
the hippocampus in schizophrenics y, 5-HTT is the
Academy of Sciences, 294 Taiyuan Road, Shanghai 200031, China. Tel.: +86 21 62822491; fax: +86 21 62822491.
primary target of SSRIs which are used as a first-line treatment for
∗∗ Corresponding author at: Institute of Neuropsychiatric Science and Systems
Biological Medicine, Changning Institute of Mental Health, the Bio-X Center Hospital,
A lot of genetic association analyses between two common poly-
299 XieHe Road, Shanghai 200042, China. Tel.: +86 21 62822491;
morphisms of 5-HTT have been carried out in different populations,
E-mail addresses: (L. He), (Y. Shi).
5-HTTLPR is a 44-bp insertion/deletion polymorphism in the 5 reg-
1 These authors contributed equally to this work.
ulatory region, involving two major alleles, namely ‘S’ (short) and
0304-3940/$ – see front matter 2009 Elsevier Ireland Ltd. All rights reserved. doi:
C. Lin et al. / Neuroscience Letters 453 (2009) 210–213Fig. 1. Diagram of the genomic structure of 5-HTT gene and the positions of the markers studied in the present work.
‘L’ (long) allele. STin2 VNTR is a 17-bp variable-number tandem-
atric Association). The diagnosis was checked and verified by two
repeats (VNTR) polymorphism, located in intron 2 and involving
independent senior psychiatrists who reviewed the psychiatric case
two major alleles, STin2.10 and STin2.12, which respectively cor-
records. Written informed consent was obtained from either the
respond to 10- and 12-repeat units of the 17-bp VNTR. Additional
participants or participants’ relatives, after the procedure had been
low frequency alleles have been identified including 7- and 9-repeat
units. Functional studies on this polymorphism have shown that the
Beside STin2, we included SNPs from the dbSNP database
amount of 5-HTT protein is significantly higher in cells carrying the
12-repeat allele when activated by morphogens in devel-
oping mouse brains STin2 acts as a transcriptional regulator in an
Haploview ver. 2.05 software ee intronic and non-functional
allele-dependent way many studies on the association
SNPs, rs2054847, rs140700 and rs2020942 were expected to belong
between 5-HTTLPR and STin2 and schizophrenia were reported, the
to the STin2-containing haplotype block oreover, tagger SNP
results have been conflicting recent progress
analysis by Haploview showed they were among the tagger SNPs
is a meta-analysis on these two polymorphisms finding signifi-
of this block. Thus these three SNPs were chosen for the present
cant association between STin2 VNTR and schizophrenia, while
study. The positions of the chosen markers were shown in
5-HTTLPR polymorphism showed no association more recent
Genomic DNA was prepared from venous blood using the stan-
study failed to detect positive association between STin2 VNTR and
dard phenol/chloroform method. For genotyping of STin2 VNTR,
schizophrenia, but it showed that the haplotype consisting STin2
primers were: forward: 5 -FAM- TCAGTATCACAGGCTGCGAG-3 ;
VNTR was associated with schizophrenia, while 5-HTTLPR did not
reverse: 5 - TGTTCCTAGTCTTACGCCAGTG-3 . PCR were conducted
belong to the same haplotype block efore, the haplotype
in a final volume of 5 l consisting of 10 ng DNA, 0.1 l 10 M
block containing STin2 VNTR emerges as a candidate for risk factor
primers, and 1 U HotStart Taq polymerase (Qiagen). MegaBACE
1000 capillary electrophoresis (Amersham Biosciences, Piscataway,
On the other hand, the associations between single nucleotide
NJ) were used to discern the genotype of 10- and 12-repeat of the
polymorphisms (SNPs) of 5-HTT and schizophrenia have been much
less explored compared with STin2 VNTR and 5-HTTLPR.
SNP genotyping was performed using ligase detection reaction.
In the present study, we attempt to evaluate the role of 5-HTT
Fluorescence-labeled primer was employed to detect the genotype
in the pathogenesis of schizophrenia using a case-control approach
of the (GT)n repeat using MegaBACE 1000 capillary electrophoresis.
involving 329 patients and 288 healthy subjects drawn from the
CLUMP 2.2 was used to compare the allelic, genotypic fre-
Han Chinese population. STin2 VNTR and three SNPs of 5-HTT gene
quencies and haplotype distribution between cases and controls
(rs2054847, rs140700, and rs2020942) were investigated.
erroni correction was used for multiple testing. Analysis
329 unrelated schizophrenic patients (203 males, 126 females,
of Hardy–Weinberg equilibrium and linkage disequilibrium were
mean age 49.54 ± 11.48) and 288 unrelated healthy subjects (159
males, 129 females, mean age 37.02 ± 8.16) were recruited in this
study. All the subjects were from Shanghai and were Han Chinese in
All genotype frequencies of the three SNPs and STin2 VNTR were
origin. All of the controls were interviewed to exclude any history
in Hardy–Weinberg equilibrium (P > 0.05, We observed no
of psychiatric disorder. Subjects with schizophrenia were strictly
significant deviation in allelic or genotypic frequencies of any single
diagnosed according to the criteria of DSM-IV (American Psychi-
one for the four markers examined For the allele frequen-
Table 1 Allelic and genotypic frequencies of four markers among schizophrenic patients and normal controls.
a Chi-square values or empirical P-values obtained using CLUMP (10,000 simulation).
c Test for departure from Hardy–Weinberg expectation in case and control groups using SHEsis. C. Lin et al. / Neuroscience Letters 453 (2009) 210–213
with schizophrenia, suggesting that the 5-HTT region contributing
Estimation of linkage disequilibrium among the four markers.
genetic risk to schizophrenia lay in the haplotype block containing
STin2. This also strengthened the data of the meta-analysis
In this study, we did not detect statistically significant associ-
ation between STin2 VNTR and schizophrenia (P = 0.07) as did the
meta-analysis, and this suggests that some allele combinations may
constitute a risk genetic background, rather than any individual
allele. Considering the P-value was <0.1% and the power was 66.0%at OR = 1.5, however, it could not be concluded that the lack of sig-nificant association is a negative result. It should be noted that a
cies, the P-value was 0.94 and the odds ratio (OR) was 0.99 for both
trend of increase in STin2 allele 12 frequencies was observed in
rs2020942 and rs2054847; the P-value was 0.18 and the OR was
schizophrenic patients tent with the result of the meta-
0.68 for rs140700; the P-value was 0.07 and the OR was 1.48 for
analysis. This is also reminiscent of the studies of Liu et al. and
STin2 VNTR. As to the genotype frequencies, the P-value of both
Tsai et al. which reported marginal association of STin2 VNTR with
rs2020942 and rs2054847 was 1.00, the P-value for rs140700 was
schizophrenia in Chinese population, with an odds ratio of approxi-
0.42, and the P-value for STin2 VNTR was 0.10. The power of both
mately 2.0 odds ratio of STin2 VNTR in our study is 1.48,
rs2054847 and rs140700 was 78.9%, power of rs140700 was 30.8%,
and this discrepancy may be also due to difference in sample size.
and power of STin2 VNTR was 66.0% at the level of OR = 1.5.
Thus, a larger sample size may be required for a conclusive result on
ws the pair-wise linkage disequilibrium (LD) among
association between STin2 VNTR and schizophrenia. Finally, while
each pair of markers. The three SNPs are in strong LD (D ≥ 0.7),
the meta-analysis reported increased STin2 allele 12 frequencies in
and in particular rs2020942 and rs2054847 were in complete LD
schizophrenic patients, our study found two haplotypes were asso-
(D = 1.0, R2 = 1.0), which is inconsistent to the genotype data in
ciated with schizophrenia, the one containing allele 12 (P = 0.011)
HapMap database. Whereas LD between SNPs and microsatellites
and the one containing STin2 allele 10 (P = 5.38 × 10−5). Since haplo-
was much weaker (D = 0.571 and 0.29).
type analysis has taken into consideration the frequencies of single
A three-marker haplotype analysis was conducted with
alleles, the difference between single allele and haplotype together
rs2054847, rs140700 and STin2 VNTR, since rs2020942 was in
with different sample size may result in this discrepancy.
total LD with rs2054847. wed the estimated haplotype
rs2054847 and rs2020942 turned out to be in total LD, which was
frequencies in both groups, with Bonferroni corrected P-values.
inconsistent with the LD data in the HapMap database, suggesting
Globally, P = 4.67 × 10−6 after Bonferroni correction, indicating a
they belonged to one big LD block which may span across 3 region
significant association. The haplotype A-G-10 (#4 in
of 5-HTT gene including STin2 VNTR. But the LD between these
displayed a strong association with schizophrenia (Bonferroni cor-
two SNPs with STin2 VNTR was not as high as expected, although
rected P = 5.38 × 10−5), carried by about 8.4% of the schizophrenic
rs2020942 is localized near to STin2 polymorphism (and
patients versus 1.8% of controls. In addition, The 5# haplotype A-
rs2054847 was predicted to be in the same LD block with STin2
G-12 is also significantly associated with schizophrenia (Bonferroni
corrected P-value = 0.011), carried by about 69.9% of patients versus
One recent neurochemical hypothesis on the etiology of
schizophrenia has implicated a primary cortical hypoglutamater-
No other significant associations were observed.
gia that enhances subcortical hyperdopaminergia and cortical
In this study, we carried out a case-control study of the associ-
hypodopaminergia In our view, 5-HT may contribute signif-
ation between four markers in 5-HTT and schizophrenia based on
icantly to the circuit alteration due to its extensive interaction
subjects drawn from the Han Chinese population. While no single
with other neurotransmitter systems especially the glutamate sys-
locus associations was observed, we found that the frequencies of
tem. The physiological basis for this view is the role of 5-HT
two haplotypes were significantly different between the patients
in the extensive biological processes which are relevant to the
positive schizophrenic symptoms (such as hallucination), the neg-
Our results support 5-HTT as a risk factor for schizophrenia in
ative symptoms (such as diminished emotional expression), and
the Han Chinese population. This is consistent with some but not
the cognitive symptoms (such as impaired long-term memories).
all previous reports. A study on a Japanese cohort investigating four
Importantly, 5-HT is also involved in the early brain development
SNPs of 5-HTT as well as 5-HTTLPR and 5-HTTVNTR, found no asso-
5-HT neuronal network is among the earliest to develop
ciation with schizophrenia However, a meta-analysis found
in embryogenesis, where 5-HT may influence neurogenesis, neu-
highly significant association between STin2 VNTR and schizophre-
ronal apoptosis, and synaptic plasticity The involvement of
nia from 12 population-based association studies hat is more,
5-HT system components in the therapeutic action of antipsychotic
a recent association study by Zaboli et al. on Caucasians revealed
drugs supports a role of 5-HT system in pathogenesis of schizophre-
a significant association between a haplotype consisting of STin2
nia Consistently, other components of 5-HT system in
VNTR and schizophrenia erestingly, we repeated the result
addition to 5-HTT were also reported to be in association with
of Zaboli et al. that the STin2-containing haplotype was associated
schizophrenia The critical role of 5-HTT in regulating 5-HT
Table 3 Estimated haplotype frequencies in the case-control subjects. 0.00000538 P = 0.000000467
a Haplotypes with a frequency <3% in both group are not shown.
b P-value Bonferroni corrected for multiple testing. Significant P-values (<0.05) are in boldface. C. Lin et al. / Neuroscience Letters 453 (2009) 210–213
transmission makes it a promising candidate gene for schizophre-
[17] W. Liu, N. Gu, G. Feng, S. Li, S. Bai, J. Zhang, T. Shen, H. Xue, G. Breen, D. St Clair, L.
nia. And our results have added a support to the role of 5-HT
He, Tentative association of the serotonin transporter with schizophrenia andunipolar depression but not with bipolar disorder in Han Chinese, Pharmaco-
transmission in the pathogenesis of schizophrenia.
In conclusion, here we conducted a case-control association
[18] J. Liu, S.H. Juo, A. Dewan, A. Grunn, X. Tong, M. Brito, N. Park, J.E. Loth, K. Kanyas,
study between genetic polymorphisms in 5-HTT and schizophre-
B. Lerer, J. Endicott, G. Penchaszadeh, J.A. Knowles, J. Ott, T.C. Gilliam, M. Baron,Evidence for a putative bipolar disorder locus on 2p13–16 and other potential
nia in the Han Chinese population. We demonstrated association
loci on 4q31, 7q34, 8q13, 9q31, 10q21–24, 13q32, 14q21 and 17q11-12, Mol.
between two STin2-containing haplotype combinations of the
5-HTT gene and schizophrenia. Therefore, we provide further evi-
[19] A. MacKenzie, J. Quinn, A serotonin transporter gene intron 2 polymorphic
dence for a role of 5-HTT in pathogenesis of schizophrenia.
region, correlated with affective disorders, has allele-dependent differentialenhancer-like properties in the mouse embryo, Proc. Natl. Acad. Sci. U.S.A. 96(1999) 15251–15255. Acknowledgements
[20] H.Y. Meltzer, Role of serotonin in the action of atypical antipsychotic drugs, Clin.
[21] L. Naylor, B. Dean, K. Opeskin, G. Pavey, C. Hill, N. Keks, D. Copolov,
We would sincerely thank all the subjects for their par-
Changes in the serotonin transporter in the hippocampus of subjects with
ticipation in this study and all the medical staff involved in
schizophrenia identified using [3H]paroxetine, J. Neural. Transm. 103 (1996)
sample-collection and diagnosis. The study was supported by the
[22] M.J. Owen, N. Craddock, M.C. O’Donovan, Schizophrenia: genes at last? Trends
Knowledge Innovation Program of the Chinese Academy of Sci-
ences, grant no. KSCX2-YW-R-01, the National 863 Program of
[23] M.J. Owens, C.B. Nemeroff, Role of serotonin in the pathophysiology of depres-
China (no.2006AA02A407), Shanghai Leading Academic Discipline
sion: focus on the serotonin transporter, Clin. Chem. 40 (1994) 288–295.
[24] S. Ramamoorthy, A.L. Bauman, K.R. Moore, H. Han, T. Yang-Feng, A.S. Chang,
V. Ganapathy, R.D. Blakely, Antidepressant- and cocaine-sensitive humanserotonin transporter: molecular cloning, expression, and chromosomal local-
References
ization, Proc. Natl. Acad. Sci. U.S.A. 90 (1993) 2542–2546.
[25] S.C. Risch, C.B. Nemeroff, Neurochemical alterations of serotonergic neuronal
systems in depression, J. Clin. Psychiatry (53 Suppl.) (1992) 3–7.
[1] A. Abi-Dargham, Alterations of serotonin transmission in schizophrenia, Int.
[26] C.A. Ross, R.L. Margolis, S.A. Reading, M. Pletnikov, J.T. Coyle, Neurobiology of
Rev. Neurobiol. 78 (2007) 133–164.
schizophrenia, Neuron 52 (2006) 139–153.
[2] G.K. Aghajanian, G.J. Marek, Serotonin model of schizophrenia: emerging role
[27] P.A. Saiz, M.P. Garcia-Portilla, C. Arango, B. Morales, V. Alvarez, E. Coto, J.M.
of glutamate mechanisms, Brain Res. Brain Res. Rev. 31 (2000) 302–312.
Fernandez, M.T. Bascaran, M. Bousono, J. Bobes, Association study of serotonin
[3] J.A. Badner, E.S. Gershon, Meta-analysis of whole-genome linkage scans of bipo-
2A receptor (5-HT2A) and serotonin transporter (5-HTT) gene polymorphisms
lar disorder and schizophrenia, Mol. Psychiatry 7 (2002) 405–411.
with schizophrenia, Prog. Neuropsychopharmacol. Biol. Psychiatry 31 (2007)
[4] J.C. Barrett, B. Fry, J. Maller, M.J. Daly, Haploview: analysis and visualization of
LD and haplotype maps, Bioinformatics 21 (2005) 263–265.
[28] A. Schotte, P.F. Janssen, W. Gommeren, W.H. Luyten, P. Van Gompel, A.S. Lesage,
[5] A. Breier, Serotonin, schizophrenia and antipsychotic drug action, Schizophr.
K. De Loore, J.E. Leysen, Risperidone compared with new and reference antipsy-
chotic drugs: in vitro and in vivo receptor binding, Psychopharmacology (Berl.)
[6] B. Dean, K. Opeskin, G. Pavey, L. Naylor, C. Hill, N. Keks, D.L. Copolov,
[3H]paroxetine binding is altered in the hippocampus but not the frontal cortex
[29] A. Serretti, R. Lilli, C. Lorenzi, E. Lattuada, C. Cusin, E. Smeraldi, Serotonin
or caudate nucleus from subjects with schizophrenia, J. Neurochem. 64 (1995)
transporter gene (5-HTTLPR) and major psychoses, Mol. Psychiatry 7 (2002)
[7] E. Erdfelder, F. Faul, A. Buchner, GPOWER: a general power analysis program,
[30] P.C. Sham, D. Curtis, Monte Carlo tests for associations between disease and
Behav. Res. Methods Instrum. Comput. 28 (1996) 1–11.
alleles at highly polymorphic loci, Ann. Hum. Genet. 59 (1995) 97–105.
[8] J.B. Fan, P. Sklar, Meta-analysis reveals association between serotonin trans-
[31] Y.Y. Shi, L. He, SHEsis, a powerful software platform for analyses of linkage dis-
porter gene STin2 VNTR polymorphism and schizophrenia, Mol. Psychiatry 10
equilibrium, haplotype construction, and genetic association at polymorphism
[9] C.E. Fiskerstrand, E.A. Lovejoy, J.P. Quinn, An intronic polymorphic domain often
[32] M.S. Sodhi, E. Sanders-Bush, Serotonin and brain development, Int. Rev. Neuro-
associated with susceptibility to affective disorders has allele dependent differ-
ential enhancer activity in embryonic stem cells, FEBS Lett. 458 (1999) 171–174.
[33] P.F. Sullivan, K.S. Kendler, M.C. Neale, Schizophrenia as a complex trait: evi-
[10] E.V. Gurevich, J.N. Joyce, Alterations in the cortical serotonergic system in
dence from a meta-analysis of twin studies, Arch. Gen. Psychiatry 60 (2003)
schizophrenia: a postmortem study, Biol. Psychiatry 42 (1997) 529–545.
[11] M.K. Hahn, R.D. Blakely, The functional impact of SLC6 transporter genetic vari-
[34] T. Sumiyoshi, C.A. Stockmeier, J.C. Overholser, G.E. Dilley, H.Y. Meltzer,
ation, Annu. Rev. Pharmacol. Toxicol. 47 (2007) 401–441.
Serotonin1A receptors are increased in postmortem prefrontal cortex in
[12] B.P. Sokolov, I. Hemandez, Abnormal expression of serotonin transporter mRNA
schizophrenia, Brain Res. 708 (1996) 209–214.
in the frontal and temporal cortex of schizophrenics, Mol. Psychiatry 2 (1997)
[35] S.J. Tsai, W.C. Ouyang, C.J. Hong, Association for serotonin transporter gene
variable number tandem repeat polymorphism and schizophrenic disorders,
[13] M. Ikeda, N. Iwata, T. Suzuki, T. Kitajima, Y. Yamanouchi, Y. Kinoshita, N. Ozaki,
Neuropsychobiology 45 (2002) 131–133.
No association of serotonin transporter gene (SLC6A4) with schizophrenia and
[36] C. Vaquero Lorenzo, E. Baca-Garcia, M. Diaz-Hernandez, C. Botillo-Martin,
bipolar disorder in Japanese patients: association analysis based on linkage
M.M. Perez-Rodriguez, C. Fernandez-Ramos, M.D. Saiz-Gonzalez, F.J. Quintero-
disequilibrium, J. Neural. Transm. 113 (2006) 899–905.
Gutierrez, J. Saiz-Ruiz, J. Fernandez Piqueras, J.L. Gonzalez de Rivera, J. de
[14] J.N. Joyce, A. Shane, N. Lexow, A. Winokur, M.F. Casanova, J.E. Kleinman, Sero-
Leon, Association between the T102C polymorphism of the serotonin-2A recep-
tonin uptake sites and serotonin receptors are altered in the limbic system of
tor gene and schizophrenia, Prog. Neuropsychopharmacol. Biol. Psychiatry 30
schizophrenics, Neuropsychopharmacology 8 (1993) 315–336.
[15] M. Laruelle, L.S. Kegeles, A. Abi-Dargham, Glutamate, dopamine, and
[37] G. Zaboli, E.G. Jonsson, R. Gizatullin, A. De Franciscis, M. Asberg, R. Leopardi,
schizophrenia: from pathophysiology to treatment, Ann. N.Y. Acad. Sci. 1003
Haplotype analysis confirms association of the serotonin transporter (5-HTT)
gene with schizophrenia but not with major depression, Am. J. Med. Genet. B
[16] J.A. Lieberman, R.B. Mailman, G. Duncan, L. Sikich, M. Chakos, D.E. Nichols, J.E.
Neuropsychiatr. Genet. 147 (2008) 301–307.
Kraus, Serotonergic basis of antipsychotic drug effects in schizophrenia, Biol. Psychiatry 44 (1998) 1099–1117.
Escritos del paciente. EL ESKAPE DEL PANÓPTICO. "Psicosis fuera del sistema" Habitación de contención. Una cama con base metálica. El metal está corroído. La habitación es pequeña y no tiene ni aberturas ni ventana. Todo es color amarillo desgastado. La puerta es de metal gris y está toda arañada. En una esquina hay un inodoro blanco sin tapa. Y una canilla de agua

 Neuroscience Letters 453 (2009) 210–213
Haplotype analysis confirms association of the serotonin transporter (5-HTT)gene with schizophrenia in the Han Chinese population
Chuwen Lin , Wei Tang , Jianxin Hu , Linhan Gao , Ke Huang , Yifeng Xu ,Guang He , Peiji Liang , Guoyin Feng , Lin He , Yongyong Shi
a Bio-X Center, Shanghai Jiao Tong University, Shanghai, Chinab Institute of Neuropsychiatric Science and Systems Biological Medicine, Shanghai Jiao Tong University, Shanghai, Chinac Changning Institute of Mental Health, the Bio-X Center Hospital, Shanghai, Chinad Institute for Nutritional Sciences, SIBS, Chinese Academy of Sciences, Shanghai, Chinae Shanghai Institute of Mental Health, Shanghai, Chinaf School of Life Science and Technology, Shanghai Jiao Tong University, Shanghai, China
Serotonin transmission has long been suspected as being involved in the pathogenesis of schizophre-
nia. 5-HTT is a promising candidate gene for schizophrenia due to its critical role in regulating serotonin
Received in revised form 10 February 2009
transmission and role in the mechanism of the atypical antipsychotic drugs. A common polymorphism
STin2 VNTR in the 5-HTT gene has been extensively investigated in the genetic association studies, butthe results are conflicting. Meanwhile, the SNPs of the 5-HTT gene have been much less explored. We
therefore conducted a case-control study of the association between STin2 VNTR and three tagging
SNPs in 5-HTT and schizophrenia in the Han Chinese population based on a cohort of 329 schizophrenic
patients and 288 control subjects. No association was found in the single locus, but haplotype-based anal-
yses revealed significant association between two haplotypes with schizophrenia even after Bonferroni
correction (P = 0.00000538 and 0.011).
Neuroscience Letters 453 (2009) 210–213
Haplotype analysis confirms association of the serotonin transporter (5-HTT)gene with schizophrenia in the Han Chinese population
Chuwen Lin , Wei Tang , Jianxin Hu , Linhan Gao , Ke Huang , Yifeng Xu ,Guang He , Peiji Liang , Guoyin Feng , Lin He , Yongyong Shi
a Bio-X Center, Shanghai Jiao Tong University, Shanghai, Chinab Institute of Neuropsychiatric Science and Systems Biological Medicine, Shanghai Jiao Tong University, Shanghai, Chinac Changning Institute of Mental Health, the Bio-X Center Hospital, Shanghai, Chinad Institute for Nutritional Sciences, SIBS, Chinese Academy of Sciences, Shanghai, Chinae Shanghai Institute of Mental Health, Shanghai, Chinaf School of Life Science and Technology, Shanghai Jiao Tong University, Shanghai, China
Serotonin transmission has long been suspected as being involved in the pathogenesis of schizophre-
nia. 5-HTT is a promising candidate gene for schizophrenia due to its critical role in regulating serotonin
Received in revised form 10 February 2009
transmission and role in the mechanism of the atypical antipsychotic drugs. A common polymorphism
STin2 VNTR in the 5-HTT gene has been extensively investigated in the genetic association studies, butthe results are conflicting. Meanwhile, the SNPs of the 5-HTT gene have been much less explored. We
therefore conducted a case-control study of the association between STin2 VNTR and three tagging
SNPs in 5-HTT and schizophrenia in the Han Chinese population based on a cohort of 329 schizophrenic
patients and 288 control subjects. No association was found in the single locus, but haplotype-based anal-
yses revealed significant association between two haplotypes with schizophrenia even after Bonferroni
correction (P = 0.00000538 and 0.011).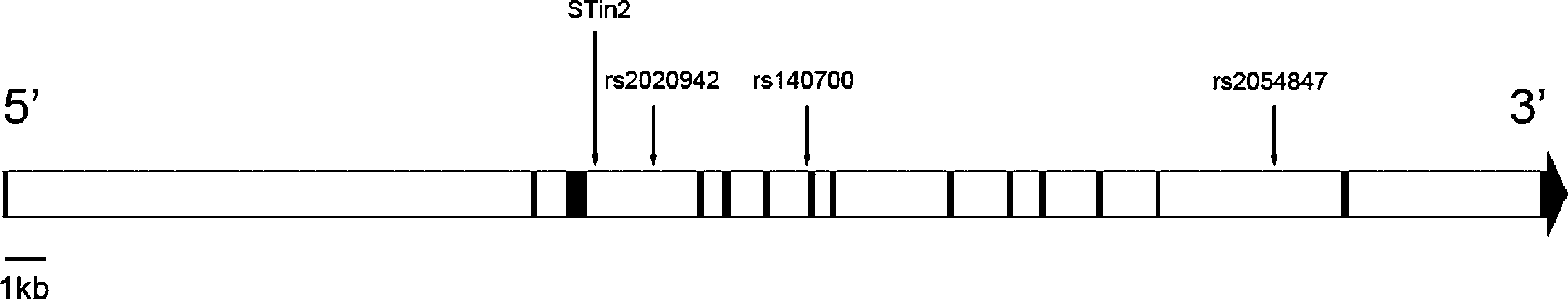 C. Lin et al. / Neuroscience Letters 453 (2009) 210–213
Fig. 1. Diagram of the genomic structure of 5-HTT gene and the positions of the markers studied in the present work.
C. Lin et al. / Neuroscience Letters 453 (2009) 210–213
Fig. 1. Diagram of the genomic structure of 5-HTT gene and the positions of the markers studied in the present work.