A New Generation of Melan-A/MART-1 Peptides That Fulfill Both Increased Immunogenicity and High Resistance to Biodegradation: Implication for Molecular Anti-Melanoma Immunotherapy1 Jean-Se´bastien Blanchet,* Danila Valmori,† Isabelle Dufau,* Maha Ayyoub,† Christophe Nguyen,* Philippe Guillaume,† Bernard Monsarrat,* Jean-Charles Cerottini,† Pedro Romero,† and Jean Edouard Gairin2* Intense efforts of research are made for developing antitumor vaccines that stimulate T cell-mediated immunity. Tumor cells specifically express at their surfaces antigenic peptides presented by MHC class I and recognized by CTL. Tumor antigenic peptides hold promise for the development of novel cancer immunotherapies. However, peptide-based vaccines face two major limitations: the weak immunogenicity of tumor Ags and their low metabolic stability in biological fluids. These two hurdles, for which separate solutions exist, must, however, be solved simultaneously for developing improved vaccines. Unfortunately, attempts made to combine increased immunogenicity and stability of tumor Ags have failed until now. Here we report the successful design of synthetic derivatives of the human tumor Ag Melan-A/MART-1 that combine for the first time both higher immunogenicity and high peptidase resistance. A series of 36 nonnatural peptide derivatives was rationally designed on the basis of knowledge of the mechanism of degradation of Melan-A peptides in human serum and synthesized. Eight of them were efficiently protected against proteolysis and retained the antigenic properties of the parental peptide. Three of the eight analogs were twice as potent as the parental peptide in stimulating in vitro Melan-specific CTL responses in PBMC from normal donors. We isolated these CTL by tetramer-guided cell sorting and expanded them in vitro. The resulting CTL efficiently lysed tumor cells expressing Melan-A Ag. These Melan-A/MART-1 Ag derivatives should be considered as a new generation of potential immunogens in the development of molecular anti-melanoma vaccines. The Journal of Immunology, 2001, 167: 5852–5861.
T hemolecularcharacterizationoftumorAgsthatcanbe BecauseofthehighfrequencyofMelan-Acirculatingprecur-
recognized by CTL has opened new opportunities for the
sors (5, 7) and evidence of strong immune responses to this Ag in
development of Ag-specific cancer vaccines (1, 2). In the
melanoma patients (8), Melan-A represents an attractive candidate
case of melanoma, a candidate Ag is Melan-A/MART-1 (Melan-A
for generic immunotherapy of HLA-A*0201ϩ melanoma patients.
hereafter), a melanocyte lineage-specific protein expressed in
A superagonist variant of the nonameric Melan-A
ϳ75–100% of primary and metastatic melanomas depending on
been shown to elicit an enhanced anti-melanoma CD8ϩ CTL re-
their clinical stage (3). Melan-A-specific CTL have been fre-
sponse (9). We have recently undertaken clinical trials of peptide
quently isolated from PBMC of both HLA-A*0201 healthy donors
vaccination using the decameric analog Melan-A
and melanoma patients and from tumor-infiltrating lymphocytes of
(ELAGIGILTV). In addition to exhibiting improved HLA-A*0201
melanoma patients (4, 5). These CTL have been shown to primar-
binding properties (higher affinity and more stable HLA-A*0201/
tigenicity and immunogenicity than the natural Melan-A peptides(10, 11). Moreover, the large majority of CTL raised either in vitroor in vivo against Melan-A
*Laboratoire d’ImmunoPharmacologie Structurale, Institut de Pharmacologie et Bi-
the Melan-A parental peptide sequences and able to specifically
ologie Structurale, Centre National de la Recherche Scientifique, Toulouse, France;
lyse Melan-A-expressing tumor cells (10 –12).
and †Ludwig Institute for Cancer Research, Division of Clinical Onco-Immunology,University Hospital and Lausanne Branch, Lausanne, Switzerland
When compared with other vaccination strategies (13), the use
Received for publication July 20, 2001. Accepted for publication September 18, 2001.
of antigenic peptides derived from tumor Ags (tumor Ag-derived
The costs of publication of this article were defrayed in part by the payment of page
peptides) as immunogens offers a number of advantages, including
charges. This article must therefore be hereby marked advertisement in accordance
low cost and facility of administration of the vaccine, high spec-
with 18 U.S.C. Section 1734 solely to indicate this fact.
ificity of elicited immune responses, and low toxicity (14, 15).
1 This work was supported in part by grants from the Center National de la Recherche
However, the use of tumor Ag-derived peptides for cancer immu-
Scientifique, the Association pour la Recherche sur le Cancer (Contract 5485), and theConseil Re´gional Midi-Pyre´ne´es (Contract 99001129). J.-S.B. is the recipient of a
notherapy faces two major limitations: the weak antigenicity and
doctoral fellowship from the Association pour la Recherche sur le Cancer and a
immunogenicity of tumor Ags (16) and their high susceptibility to
European Molecular Biology Organization short-term fellowship (ASF-9575). M.A. is a recipient of a postdoctoral fellowship from the European Community-Biomed
proteolytic degradation by proteases (17–20). Weak immunoge-
nicity can partially be ascribed to a certain level of immune tol-
2 Address correspondence and reprint requests to Dr. Jean Edouard Gairin, Institut de
erance to self-derived sequences (including suboptimal MHC
Pharmacologie et de Biologie Structurale, Unité Mixte de Recherche 5089, Centre
binding and/or T cell recognition). Limiting the extent of peptide
National de la Recherche Scientifique, 205 route de Narbonne, 31400 Toulouse,France. E-mail address: [email protected]
degradation by, for example, delivering the peptide incorporated in
Copyright 2001 by The American Association of Immunologists
a water/oil emulsion often significantly increases immunogenicity
of a free amino group with an F-moc-protected preformed amino aldehyde
(21). A more effective approach for avoiding rapid peptide degra-
(31). N-terminal hydroxypeptides were synthesized following a previously
dation by proteases implies the design of protease-resistant peptide
described procedure (32). The retro-inverso bond ⌿(NH-CO) was obtainedby replacement of two sequential amino acids with an (R,S)-2-substituted
analogs. Approaches based on structural modifications to inhibit
malonate derivative and a gem-diaminoalkyl residue (for example, the
proteolytic degradation of bioactive peptides exist (22, 23) and
gem-diaminoalkyl residue corresponding to glutamic acid side chain and
have been applied to MHC class I-restricted (20, 24, 25) or class
2-substituted malonic acid corresponding to leucine side chain (2(R,S)-
II-restricted (26) antigenic peptides. Unfortunately, the structural
isobutylmalonic acid) were used for the synthesis of [⌿
A27L) (33, 34). A mixture of two diastereoisomers of the
modifications introduced in the antigenic peptide sequence most
modified peptide was obtained. The two isomeric forms were not sepa-
often result in a dramatic reduction or even in a complete loss of
rated. Peptides were purified (purity, Ն98%) by RP-HPLC on a C column
peptide binding to MHC and/or T cell recognition (24, 27–29).
(Aquapore Brownlee, PerkinElmer, Norwalk, CT). The identities of the
This difficulty may be partly overcome by a detailed knowledge of
purified peptides were confirmed by electrospray ionization-mass spec-
the degradation pathway of the antigenic peptide in human serum
trometry (ESI-MS). Peptide stock solutions were made (10Ϫ3 M in 100%DMSO) and stored at Ϫ20°C.
that allows the rational design of minimally modified, peptidase-resistant, and still biologically active nonnatural analogs (20, 24). Degradation of Melan-A peptides in human serum
It is clear that synthetic derivatives of tumor Ags combining both
Peptides were added to preheated (10 min at 37°C before the assay) human
high protection against peptidases and higher immunogenicity rep-
serum to a final concentration of ϳ0.5 ϫ 10Ϫ4 M and incubated at 37°C.
resent candidates of choice for cancer immunotherapy. However,
For the analysis of peptide persistence in serum with Melan-A-specific
none has been successfully designed and used until now.
CTL recognition assay, aliquots (100 l) were removed at different timesand put in liquid nitrogen to stop the enzymatic reaction. 51Cr-labeled T2
The aim of the present study was to design such derivatives of
cells were then pulsed with serial dilutions of each degradation sample at
the Melan-A tumor Ag. For that, we merged complementary ap-
4°C, and chromium release experiments were performed as previously de-
proaches based on manipulation of the structural and pharmaco-
logical properties of MHC class I-restricted Ags and of the im-
For on-line RP-HPLC/ESI-MS analysis, aliquots of the degradation so-
mune response against cancer. First we dissected the mechanism of
lution (100 l) were removed at different times, and the enzymatic reactionwas stopped by addition of 11 l of trifluoroacetic acid (TFA). Precipitated
Melan-A peptide degradation in human serum. Second, on the ba-
serum proteins were pelleted by centrifugation at 15,000 rpm for 10 min at
sis of these results, we designed a series of 36 nonnatural deriva-
4°C. The supernatants were frozen and kept at Ϫ20°C until analysis. We
checked that 1) the Melan-A peptides did not precipitate and were stable in
them, eight were fully resistant to proteolysis by serum proteases
the presence of 10% TFA, and 2) DMSO did not affect serum proteaseactivities at the concentration used. The HPLC profile of serum alone pre-
and retained the antigenicity and immunogenicity of the parental
cipitated by 10% TFA was recorded to detect nonprecipitated peptides
peptide. When used to stimulate in vitro Melan-specific CTL re-
present in the serum. Melan-A peptides and their degradation products
sponses in PBMC from normal donors, three of these sequences
were separated, and their sequences were determined and quantified by
were more potent activators of tumor-reactive CTL than the pa-
rental peptide. These analogs represent a new generation of Melan-
Coulter, Palo Alto, CA), a Waters 600 MS chromatograph (Waters, Mil-ford, MA), and a TSQ-700 Finnigan-MAT mass spectrometer (Thermo
A/MART-1 Ag derivatives that now combine enhanced immuno-
Finnigan, San Jose, CA) as previously described (19, 24). Quantitative
genicity with proteolysis resistance. Therefore, they should be
determination of peptides and their degradation products were obtained
considered potential immunogens in the development of molecular
operating in the selected ion monitoring mode. Materials and Methods
Peptide binding to HLA-A*0201 was assessed in a functional competition
assay based on inhibition of recognition of the antigenic peptide tyrosi-nase
(YMDGTMSQV) by the HLA-A*0201-restricted CTL clone
The tumor cell lines Me 290 and Me 275 were established from surgically
156/34. Various concentrations of competitor peptides (50 l) were incu-
excised melanoma metastasis from patients LAU203 and LAU50, respec-
bated with 51Cr-labeled T2 cells (50 l; 1000 cells/well) for 15 min at room
tively, and maintained in culture as previously described (10). T2, an HLA-
temperature. A suboptimal dose (1 nM) of the antigenic peptide tyrosi-
A*0201ϩ human T/B cell hybrid peptide transporter-deficient cell line
(50 l) was then added together with specific CTL (5000 cells/
(30), was cultured in DMEM/10% FCS supplemented with 0.55 mM Arg,
well; 50 l). Chromium release was measured after a 4-h incubation at
0.24 mM Asn, and 1.5 mM Gln. The tyrosinase-specific CTL clone 156/34
37°C. The concentration of each competitor peptide required to achieve
was derived after in vitro stimulation of tumor-infiltrating lymphocytes
50% inhibition of target cell lysis was then determined (IC ).
from patient LAU156 with the natural tyrosinase
MSQV) (12). The polyclonal monospecific CTL line used for analog rec-
Peptide recognition by Melan-A specific CTL
ognition experiments was induced in vitro after stimulation of PBMC from
Ag recognition was assessed using chromium release assays. Target cells
(T2 cells) were labeled with 51Cr for 1 h at 37°C and washed three times. 51
Cr-labeled target cells (1000 cells/50 l) were then added to various
concentrations of antigenic peptide (50 l) in V-bottom 96-well plates for
Peptides were synthesized by the solid phase method using F-moc chem-
15 min before addition of effector cells. A polyclonal CTL line specific for
istry and the N,NЈ-diisopropylcarbodiimide/N-hydroxyaza-benzotriazole
A27L was used as effector cells and added (5000 cells/100
coupling procedure. Peptide with  amino acids, d amino acids, ␣-meth-
l) at a defined E:T cell ratio. Chromium release was measured in 100 l
ylated amino acids (␣Meaa),3 or N-methylated amino acids (NMeaa) were
of supernatant harvested after 4 h of incubation at 37°C. The percentage of
synthesized by incorporation of the corresponding modified amino acids
specific lysis was calculated as follows: % specific lysis ϭ [(experimental
commercially available (Bachem, Budendorf, Switzerland (NMeaa); Fluka,
release Ϫ spontaneous release)/(total release Ϫ spontaneous release)] ϫ
Buchs, Switzerland ( amino acids); and Acros Organic, Noisy-Le-Grand,
100. The concentration of each peptide required to achieve 50% maximal
France (␣Meaa)). Peptides with ␣Meaa were obtained as racemics, because
lysis of target (EC ) was then determined.
␣Meaa were purchased in a racemic form. Whenever possible, stereoiso-mers were isolated by reverse phase HPLC (RP-HPLC) and named p1 and
Generation of Melan-A specific CTLp2. The reduced bond ⌿(CH -NH) was formed by the reductive alkylation
PBMC from HLA-A*0201ϩ healthy donors were isolated by centrifuga-tion in Ficoll-Paque (Pharmacia Biotech, Uppsala, Sweden). CD8ϩ lym-
phocytes were isolated using a miniMACS device (Miltenyi Biotec, Ber-
Abbreviations used in this paper: ␣Meaa, ␣-methylated amino acid; ESI-MS, elec-
gisch Gladbach, Germany). The resulting population routinely contained
trospray ionization-mass spectrometry; DC, dendritic cell; NMeaa, N-methylatedamino acid; NOHG, N-hydroxylated glycine; RP-HPLC, reverse phase HPLC; TFA,
Ͼ75% CD8ϩ T cells and was used as the responder cell population. Pu-
trifluoroacetic acid; RIC, reconstructed ion current.
rified CD8ϩ T cells were plated at 1 ϫ 106 cells/well together with 2 ϫ 106
A NEW GENERATION OF MELAN-A/MART-1 Ag DERIVATIVES
stimulator cells/well in 24-well plate in a total volume of 2 ml of Iscove’s
24-h treatment in serum (Fig. 1A), although it appeared almost
medium supplemented with 10% human serum, Asn, Arg, and Gln (com-
completely degraded after 2 h (Fig. 1B). This observation can be
plete medium) in the presence of IL-7 (10 ng/ml; R&D Systems, Oxon,
explained by a 2- to 3-log difference in the sensitivity of the assays
U.K.), IL-2 (10 U/ml; Glaxo Wellcome, Geneva, Switzerland), and stim-ulating peptide (1 M). Stimulator cells were prepared as follows. Cells
(biological vs physicochemical) as we previously noted (24). It
(2 ϫ 106) derived from the CD8Ϫ population after miniMACS CD8ϩ lym-phocyte isolation were irradiated (3000 rad) and adjusted to the appropriatevolume before addition to the CD8ϩ-enriched responder cell population. On day 7 cells were restimulated with peptide-pulsed T2 cells. T2 cellswere incubated for 2 h at 37°C in serum-free medium (X-VIVO 10; Bio-Whittaker, Walkersville, MD) with the appropriate stimulating peptide (1
M) and human  -microglobulin (3 g/ml; Sigma-Aldrich, St. Louis,
MO). Peptide-pulsed T2 cells were washed, irradiated (10,000 rad), ad-justed to the appropriate volume of complete medium supplemented withIL-7 (10 ng/ml) and IL-2 (10 U/ml), and added to the lymphocyte culture(2 ϫ 105 cells/well). Subsequent restimulations were performed weeklywith peptide-pulsed T2 cells. CTL activity was first tested at the end of thefirst restimulation using an ELISPOT assay for IFN-␥ production bypeptide-reactive CTL. Flow cytometric analysis and Melan-A tetramer-guided cellsorting of specific CD8ϩ CTL
The specificity of CTL recognition was monitored by flow cytometric anal-ysis after Melan-A
A27L HLA-A*0201 tetramer staining (5, 8). Cells
were stained with tetramers (200 ng/sample) in 20 l of PBS/2% FCS for20 min at room temperature, then 20 l of anti-CD8-FITC mAb (BD Bio-sciences, Basel, Switzerland) was added, and cells were incubated for anadditional 30 min at 4°C. Cells were washed once in the same buffer andanalyzed on a FACScan (BD Biosciences, San Jose, CA) flow cytometer. Data analysis was performed using CellQuest software (BD Biosciences). The CD8ϩtetramerϩ and CD8ϩtetramerϪ lymphocyte populations weresorted using a FACSVantage (BD Biosciences) cell sorter. After cell sort-ing the polyclonal monospecific CTL population was expanded by restimu-lation in a nonspecific fashion using PHA (1 g/ml; Sigma-Aldrich), andIL-2 (150 U/ml). Results Melan-A Ag-related peptides are rapidly degraded in human serum by amino- and dipeptidyl-carboxy-peptidases
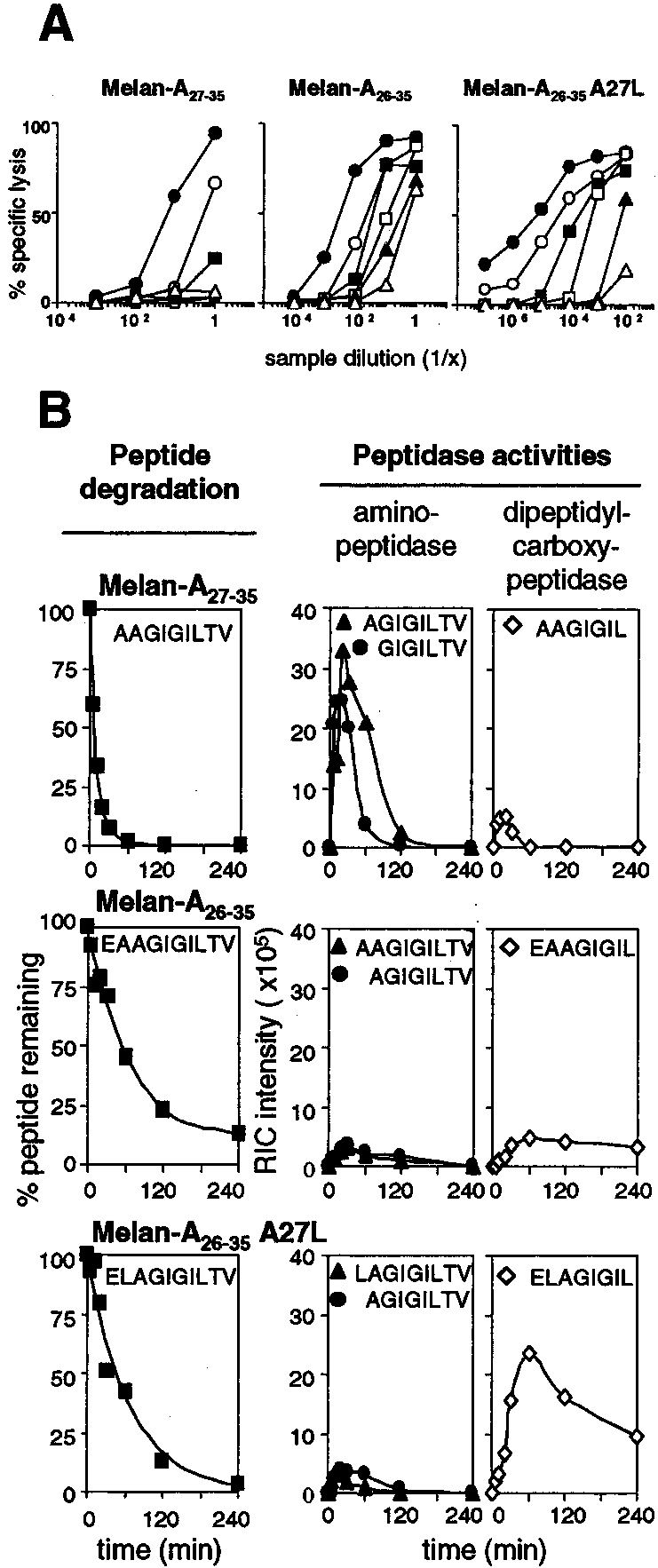
The kinetics as well as the mechanism of Melan-A peptide pro-teolytic degradation were determined by incubation of syntheticpeptides in human serum. We studied different peptides related tothe Melan-A Ag: the nonamer Melan-A
A27L (ELAGIGILTV). We first determined the persistence
of the antigenic peptide over time in serum as monitored by spe-cific CTL recognition using a chromium release assay. After dif-ferent incubation times in serum, serial dilutions of the samplescontaining the synthetic peptide were pulsed onto T2 cells andincubated with Melan-A-specific CTL. Lysis of peptide-pulsed T2cells represents the persistence of intact peptide in human serumover time. Clearly, Melan-A antigenic peptides incubated in serum
FIGURE 1.
Degradation of the Melan-A peptides in human serum. A,
very rapidly lost the ability to sensitize T2 target cells for lysis by
Melan-A peptides were incubated in human serum, and the persistence ofthe Ag was evaluated using a functional cytolytic assay. At each incubation
specific CTL (Fig. 1A), suggesting that they were degraded by
time T (F, 0 min; E, 30 min; f, 60 min; Ⅺ, 120 min; Œ, 240 min; ‚, 24 h),
peptidases in human serum. Of note, the Melan-A nonamer AA-
51Cr-labeled T2 cells were pulsed with serial dilutions of each degradation
GIGILTV seemed to be more sensitive than the two Melan-A
sample and used as target cells in the chromium release assay. The CTL
used was a polyclonal monospecific CTL line raised against Melan-A26–
The exact mechanism of antigenic peptide degradation in serum
35A27L peptide. The E:T cell ratio used was 10:1. B, Degradation kinetics
was addressed by on-line RP-HPLC/ESI-MS that allows unambig-
of Melan-A Ags analyzed by HPLC/ESI-MS. Melan-A peptides were in-
uous identification and precise quantification of the degradation
cubated in human serum, and the presence of the peptide was addressed by
products resulting from tumor Ag proteolysis (19, 24). As shown
direct detection using on-line HPLC/ESI-MS. Peptide quantification was
in Fig. 1B (left panels), the Melan-A
achieved by analysis of the mass spectrometry data. For a given molecular
, 5 min), in agreement with a previous study (20).
species, the reconstructed ion current (RIC) intensity characteristic of the
identified peptide or degradation product was determined. B, left panels,
The percentage of peptide remaining was calculated for each sample with
A27L (ELAGIGILTV) were degraded less rapidly
the amount of peptide detected at T0 as a reference and plotted against the
than the nonamer, thus confirming the initial observation, and dis-
time: (RIC intensity)T ϫ 100)/(RIC intensity)T0). B, middle and rightpanels, Kinetics of appearance of the amino- and carboxyl-terminal deg-
radation products of Melan-A Ags. The amount of each product is shown
could sensitize T2 cells for lysis by Melan-A-specific CTL after a
as the RIC intensity plotted against time.
further suggests that few copies of Melan-A
Protection against amino-peptidases was obtained by substitution
able by MS remained in the medium and were still able to stim-
of glutamic acid residue at position 1 (E1) by aa (E1, A1, or
ulate, even though very weakly, the Melan-A-specific CTL.
D1) (35), NMeaa (NMeE1), a pyro-glutamic acid (pE1), N-acety-
Identification of the amino- and carboxyl-terminal fragments al-
lated E, or an N-hydroxylated glycine (NOHG1) (32). Modifica-
lowed us to determine the mechanism of peptide degradation. The
tion of leucine residue at position 2 (L2) was achieved by replacing
degradation process of the Melan-A peptides in human serum fol-
it with methylated aa (␣MeL2, NMeL2) or dL2. To prevent deg-
lowed two main pathways involving amino- and dipeptidyl-car-
radation by dipeptidyl-carboxy-peptidase we replaced L8 by ␣MeL8
boxy-peptidase activities (Fig. 1B, middle and right panels, and
or dL8 or T9 by NMeT9 or dT9. To avoid carboxy-peptidase degra-
Fig. 2), as previously observed for the MAGE-1.A1 tumor Ag
dation, we also synthesized carboxy-amidated (CO-NH ) peptide. Fi-
(24). Degradation experiments using selective peptidase inhibitor
nally, peptide bond alterations, such as reduced bond ⌿(CH -NH) or
bestatine (amino-peptidase inhibitor) or captopril (dipeptidyl-car-
retro-inverso bond ⌿(NH-CO), were included to prevent amino- or
boxy-peptidase inhibitor) confirmed this. In both cases peptide
carboxyl-terminal degradation. All the analogs are listed in Table I.
degradation was partially inhibited (data not shown). Thus, the first
As exemplified in Fig. 3 for analogs [D1]-Melan-A and [⌿
two peptide bonds to be sensitive to peptidases are Glu1-Leu2 (or
NH)]-Melan-A, the amino- and carboxyl-terminal structural mod-
Ala2) and Leu8-Thr9. On-line RP-HPLC/ESI-MS analysis also
ifications of the Melan-A Ag protected the peptidase-sensitive
gave us an estimation of the quantity of a given degradation prod-
bond. However, one-site protection did not result in peptides with
uct present during the time course of degradation. As shown in Fig.
significantly improved stability, indicating that protection at both
1B, the main amino-terminal degradation products of the Melan-
ends is needed to stabilize the peptide. Indeed, all the two-site
nonamer (AGIGILTV and GIGILTV) were more abundant
protected analogs were rendered resistant to peptidase activities
values varying from Ͼ10 to ϾϾ24 h, as exemplified in
Fig. 3 (analog [D1, ␣MeL8]-Melan-A) and summarized in Table
amino-peptidases than the Melan-A decamers. This sensitivitycould explain the very short half-life of the nonameric peptide.
I. Among the doubly protected analogs, we noted that those bear-
Analysis of carboxyl-terminal degradation of the Melan-A pep-
ing either amidation or ␣-methylation at the C terminus displayed
tides also revealed differences between the nonamer and the
the shortest or longest half-lives, respectively.
decamers. The first carboxyl-terminal product from Melan-A27–35degradation (AAGIGIL) was detected very rapidly, peaked at 20
Nonnatural Melan-A derivatives are efficiently recognized by
min, and was totally degraded within 60 min. In the case of the two
decamers, the quantity of the first carboxyl-terminal degradationproducts (EAAGIGIL or ELAGIGIL) was maximal at 60 min.
We and others have previously shown that structural modification
These products were then degraded, but were still present in a
of an antigenic peptide may considerably affect the efficiency of
detectable quantity at 240 min. The scheme depicted in Fig. 2
peptide-MHC binding and/or CTL recognition (24, 25, 27). We
summarizes the degradation mechanism of Melan-A
first determined the HLA-A*0201 binding properties of the 36
The slower rate of degradation may reflect that glutamic acid at the
newly designed nonnatural analogs in a functional competition as-
amino terminus of the peptide is not optimal for amino-peptidase
say (10). As reported in Table I, a dramatic reduction in peptide-
activity. As a consequence, these peptides were less susceptible to
MHC binding was observed for about half the peptides. However,
amino-terminal degradation and were more available for carboxy-
for the other half, the peptide affinity for HLA-A*0201 was either
peptidase activity. Altogether, the degradation data suggest that the
nature of the amino-terminal residues of peptide has a direct or
reduced. Indeed, the structural alteration of the anchor residue L2
indirect effect on both amino- and carboxy-peptidase activities.
was often deleterious on MHC binding ([dL2]- or [NMeL2]-Melan-A with an IC
Designing synthetic antigenic peptides resistant to degradation
shown by others (27), peptide backbone alteration in reduced orretro-inverso analogs dramatically reduced the peptide affinity for
To identify Melan-A peptides resistant to proteolysis, we synthe-
MHC. Other modifications showed limited or no impact on peptide
sized a series of peptide analogs with structural modifications at
affinity for MHC even if they concerned anchor residue ([E1]-
one or both peptidase-sensitive positions and tested their degrada-
Melan-A IC , 2 nM; [␣MeL2]-Melan-A IC , 6 nM). Finally,
doubly protected Melan-A analogs such as [A1, ␣MeL8]( p1),
starting sequence because of its enhanced immunogenicity (10).
[NOHG1, ␣MeL8]( p1), or [␣MeL2, ␣MeL8] displayed HLA-A*0201 binding affinity very similar to if not identical with that ofthe parental Melan-A
Next, we tested the ability of the nonnatural Melan-A
analogs to be recognized by specific CTL (exemplified in Fig. 4and summarized in Table I). As expected, CTL recognition of thenonnatural Melan-A analogs correlated with MHC binding formost of the analogs, with poor HLA-A2 binders being weaklyrecognized by CTL. However, strikingly, most of the structurallymodified peptides were able to efficiently sensitize target cells tolysis by Melan-A-specific CTL, with EC
molar range. Interestingly, some doubly protected analogs dis-played efficiencies comparable to or even higher than that of theparental Melan-A
FIGURE 2.
man serum. Full and dotted arrows correspond to dipeptidyl-carboxy-pep-
tidases and amino-peptidases, respectively.
A NEW GENERATION OF MELAN-A/MART-1 Ag DERIVATIVES
Table I. Sequences, HLA-A*0201 binding properties, Melan-A-specific CTL recognition, and stability in human serum of Melan-A peptides andnonnatural Melan-A derivativesa Competitor activity was measured on the basis of the inhibition of recognition of the tyrosinase
(YMDGTMSQV) antigenic peptide in the context of HLA-A*0201
by the tyrosinase peptide-specific CTL clone 156/34. b Peptide half-life was determined by HPLC-ESI-MS analysis. n.t., Not tested. c Melan-A designation corresponds to Melan-A
d The values correspond to the biological activities of each stereoisomer ( p , p ) of the ␣-methylated peptide purified from resolved HPLC peaks. e The value corresponds to the biological activity of the mixture of the stereoisomers when HPLC peaks were not resolved and did not allow the separation of the stereoisomers
Selected nonnatural Melan-A analogs efficiently stimulate the
were tested by flow cytometry 7 days after the second stimulation
expansion of Melan-A-specific CTL precursors in the circulation
(MC2) for the presence of CD8ϩA2/Melan-A tetramerϩ cells. In cul-
The eight doubly substituted derivatives displaying the highest ef-
tures stimulated with the nonnatural Melan-A analogs, A2/Melan-A
ficiencies were selected for further studies. They are listed in Table II.
tetramerϩ cells were detected in the CD8ϩ cell population, indicating
To assess their immunogenicity, we used them to stimulate CD8ϩ-
that the A2/Melan-A tetramer was fully or partially cross-reactive
enriched cells isolated from PBMC. The Melan-A
with the analog-specific CTL, as shown in Fig. 5. Cross-reactivity of
was used as a positive control. For the second stimulation, irradiated
the A2/Melan-A tetramer allowed us to directly evaluate the efficacy
T2 cells pulsed with the appropriate peptide were used. All cultures
of the nonnatural analogs to generate Melan-A-specific CTLs from
FIGURE 3.
and doubly protected nonnatural Melan-A analogs. Exam-ples of stability of mono-protected ([D]LAGIGILTV:[D1]-Melan-A, left panel; ELAGIGIL-[(CH -NH)]-
(CH2-NH]-Melan-A, middle panel) and doubly
protected ([D]-LAGIGI-[␣MeL]-TV: [D1, ␣MeL8]-Melan-A, right panel) analogs in human serum areshown. Degradation was addressed using HPLC/ESI-MS detection as described for Melan-A peptides(see Fig. 1B). The amount of each peptide was deter-mined after analysis of MS data and is presented as apercentage of the maximum amount of the parentalpeptide at time zero (T0). f, Amount of peptide re-maining; F and E, amounts of degradation productsgenerated from amino- and dipeptidyl-carboxy-pepti-dase activities, respectively.
PBMC of healthy donors. The percentage of Melan-A-specific cells
out specific stimulation, the antigenic specificity of the sorted cells
was tested in a chromium release assay. As shown in Fig. 5, B and
donor to another, related to the frequency of Melan-A-reactive pre-
C, the sorted CTL specific for a nonnatural Melan-A analog were
cursors in the initial CD8ϩ population (Table II; HD224, 2.3%;
able to recognize and efficiently lyse not only T2 target cells pre-
HD410, 2.5%; HD220, 0.3%). Interestingly, three analogs were able
to induce twice as many Melan-A-specific cells superior as the pa-
cell lines Me 290 and Me 275 that naturally express the Melan-A
[A1, ␣MeL8]-, [E1, ␣MeL8]-, and [D, ␣MeL8]-Melan-A (Table
Ag on their surfaces were also recognized efficiently by nonnatural
II and Fig. 5). Thus, these three analogs are more immunogenic in
Melan-A-specific CTL. The Na8 cell line that does not express
the Melan-A gene was used as a control. It was not susceptible oronly weakly susceptible to lysis by the nonnatural Melan-A-spe-
A2/Melan-A tetramer-guided isolation and functional
cific CTL, indicating the absence of nonspecific lytic activity, but
characterization of in vitro stimulated nonnatural Melan-A
became efficiently lysed when pulsed with the Melan-A
ural peptide. In conclusion, CTL induced with the nonnatural an-
To further document the Ag specificity of CTL generated upon
alogs are fully competent to recognize the parental Melan-A pep-
stimulation with the nonnatural Melan-A analogs, we sorted the
tides and lyse tumor cells that naturally express the natural
CD8ϩA2/Melan-A tetramerϩ cells. After in vitro expansion with-
FIGURE 4.
Recognition of the nonnatural, doubly protected Melan-A analogs by Melan-A
A27L-specific CTL. Recognition of the doubly protected
analogs [A1, ␣MeL8]-Melan-A, [E1, ␣MeL8]-Melan-A, [D1, ␣MeL8]-Melan-A, and [A1, V10-CONH ]-Melan-A (F, upper panels) by CTL was
addressed in a chromium release assay using a polyclonal monospecific CTL line raised against Melan-A
A27L. The target cells were T2 cells pulsed
with increasing concentrations of peptide, and the E:T cell ratio was 10:1. The Melan-A peptides Melan-A
E26A were tested as controls (E, lower panels).
A NEW GENERATION OF MELAN-A/MART-1 Ag DERIVATIVES
Table II. In vitro immunogenicity of Melan-A
less, the use of such immunogenic peptide analogs as a vaccine
may be restricted due to their high susceptibility to protease deg-radation. Thus, the next step was to introduce structural modifica-
tions to obtain proteolysis-resistant antigenic peptides. In the
present study we first elucidated the degradation pathway of the
Melan-A antigenic peptides. Then, according to these results wedesigned structurally modified nonnatural Melan-A analogs that
were potent activators of tumor-reactive CTL.
Our results are in accord with previous findings that antigenic
peptides can be rapidly degraded in human serum by proteases (17,
[A1, ␣MeL8]-Melan-A (p ) 2
18, 20, 24). The analysis of antigenic peptide persistence in serum
[E1, ␣MeL8]-Melan-A (p ) 2
using a Melan-A-specific CTL recognition assay indicates that the
[D1, ␣MeL8]-Melan-A (p ) 2
nonamer AAGIGILTV is degraded more rapidly
than the two Melan-A decamers. This observation could be ex-
plained in part by the ability of the Melan-A
a Melan-A-specific cells (A2/ELA-tetramerϩCD8ϩ) were directly detected 7 days
CTL used in the test to recognize shorter peptides that could be
after the second stimulation by flow cytometry by costaining of the culture with
intermediate degradation products of the decamers. Indeed, the
A2/ELAtetramersPE and CD8FITC mAb (see Materials and Methods for more details).
nonapeptides AAGIGILTV and LAGIGILTV resulting from re-
Analogs displaying an efficiency higher than that of the parental Melan-A
moval of the first amino acid from Melan-A
b PBMC from three different healthy donors (HD224, HD410, HD220) were stim-
35A27L, respectively, are efficiently recognized by the Melan-A-
ulated in vitro with the indicated peptide as described in Materials and Methods.
specific CTL used in the degradation test (data not shown) (10). Thislimitation was overcome by the analytical on-line HPLC-ESI/MS ap-
Discussion
proach (19). We confirmed the initial observation indicating that the
Improvement of immunogenicity of TDAP is critical for the de-
Melan-A nonamer is degraded more rapidly than the two decamers.
velopment of efficient cancer vaccines. Introducing favorable an-
More importantly, we identified and quantified the degradation frag-
chor residue in the peptide sequence to increase peptide affinity for
ments leading to outlining the exact mechanism of Melan-A peptide
MHC is a first step to improved immunogenicity (36). Neverthe-
degradation and identification of protease-sensitive bonds within the
FIGURE 5.
Antigenic specificity of, and tumor recognition by, CTL induced with the nonnatural Melan-A analogs. After in vitro stimulation of PBMC
from healthy donor HD224 with the Melan-A peptides, the tetramerϩCD8ϩ lymphocyte population was sorted on a FACSVantage cell sorter (BDBiosciences). Cell sorting of the tetramerϩCD8ϩ population and subsequent in vitro expansion of lymphocytes were realized for all stimulations with thedifferent analogs under the same conditions. Cytolytic CTL activity was measured after in vitro expansion of the different CTL populations (see Materialsand Methods). A, Dot plot representation of cytometry analysis of culture after in vitro stimulation with Melan-A
analogs. The gate defined for cell sorting of tetramerϩCD8ϩ population is represented. B, Melan-A peptide recognition by the sorted tetramerϩCD8ϩ CTLfrom the corresponding gate shown in A (‚, Melan-A
A27L; F, Melan-A doubly modified analog). The chromium
release assay was performed with T2 cells as target cells pulsed with increasing peptide concentrations and at an E:T cell ratio of 10:1. C, Tumor cellrecognition by the sorted tetramerϩCD8ϩ CTL from the corresponding gate shown in A. A cytolytic assay was performed with 51Cr-labeled cells (Mel275and Mel290: HLA-A*0201ϩ, Melan-Aϩ; Na8: HLA-A*0201ϩ, Melan-AϪ) with increasing E:T cell ratios. f, Mel275; F, Mel290; E, Na8; ϩ, Na8 andMelan-A peptide).
peptide sequence. This step is of critical importance for introducing
servation has been reported. To further assess the immunogenicity
minimal, but efficient, structural modifications in the tumor antigenic
of the nonnatural Melan-A peptides, we performed in vitro stim-
sequence. As we previously showed for other human or murine tumor
ulation of PBMC from healthy donors with the modified peptide
antigenic peptides (19, 24), the Melan-A peptides are very sensitive to
analogs. We used A2/Melan-A tetramer to quantify the number of
exopeptidases in serum. Thus, this mechanism of peptide degradation
Melan-A-specific T cells elicited after stimulation with the pro-
involving both amino- and dipeptidyl-carboxy-peptidases seems to be
tected peptides. This analysis shows that stimulation with doubly
generic for antigenic peptide degradation in serum. In addition, be-
substituted, fully protected analogs can give higher numbers of
cause some doubly protected analogs were extremely stable in serum,
Melan-A-specific T cells compared with those obtained by stim-
we can deduce that the peptidic bonds left unprotected in the antigenic
ulation with the parental peptide Melan-A
sequence are not affected by endopeptidase activity. This finding is in
sults thus indicate that protection against proteolysis of antigenic
agreement with previous observations indicating that endopeptidases
peptides could significantly enhance in vitro immunogenicity.
are not involved in the degradation of short peptides in serum (17, 37).
They further show the unique immunological properties of these
Melan-A nonamer and decamers show marked quantitative and qual-
itative differences in their degradation properties. However, the dif-
ferences in the half-life of the Melan-A peptides could not be ex-
of Melan-A-expressing tumors, the CTL elicited by stimulation
plained solely by differences in peptide length, because the decamer
A27L efficiently cross-recognize the endog-
E26A (AAAGIGILTV) displayed a kinetic of degra-
enously expressed Melan-A sequences (10, 11). We show in the
, 15 min; data not shown) closer to that of Melan-A
present study that CTL elicited by the nonnatural Melan-A analogs
than that of the two Melan-A decamers. Thus, the nature of the first
are also cross-reactive with the parental and endogenously ex-
amino-terminal residues that greatly influence peptide susceptibility to
pressed Melan-A peptides at the tumor cell surface. We thus val-
amino-terminal degradation (37) seems to govern the kinetics of deg-
idate and extend the tumor Ag cross-reactivity properties to the
radation of the Melan-A peptides. Knowledge of the exact mechanism
of peptide degradation was indeed of critical importance for rationally
Antigenic peptide stability to protease degradation could have
and efficiently designing nonnatural Melan-A analogs resistant to pro-
important implications to elicit an efficient immune response. In
tease degradation. To obtain fully protected peptides, the two pro-
peptide-based cancer vaccines, after injection, the peptides have to
tease-sensitive bonds (E1-L2 and L8-T9) of the Melan-A
be loaded onto professional APCs such as DCs. Under activation,
sequence needed to be modified. However, the doubly modified pep-
DCs overexpress not only MHC, CD40, and CD80, but also CD13
tides did not show equal resistance to proteolysis. In particular, C-
and CD26, molecules that are amino- and dipeptidyl-peptidases,
terminal amidated analogs showed the weakest resistance and were
respectively (40 – 42). Induction of such ectopeptidases on the sur-
about twice as unstable as ␣-methylated ones. We may assume that
face of activated DCs could increase the extracellular degradation
this moderate protective effect is due to the capacity of a dipeptidyl-
process of exogenous antigenic peptide and reduce the peptide
carboxy-peptidase, such as angiotensin-converting enzyme, to cleave
loading onto MHC molecules. If peptides are degraded before pre-
C-terminal amidated substrates (38). Because studies have shown that
sentation by DCs, repeated administration and/or high doses of
peptide stability is similar in FCS and in the mouse (18, 37, 39), we
antigenic peptide are then required for immunization (43, 44).
may reasonably postulate that the doubly protected analogs resistant
Thus, the effective dose of a degradable peptide cannot be exactly
to human serum proteases would also be protected against proteolysis
estimated from the dose injected. Yet the concentration of peptide
in vivo. However, in most antitumor vaccination protocols, the anti-
used to elicit efficient antitumor-specific CTL is a critical param-
genic peptide is delivered either by direct s.c. injection or after pulsing
eter (45). Indeed, high-avidity CTL, which are highly efficient for
onto dendritic cells (DCs). The biological significance of studying
antitumor response, were only induced in vitro and in vivo with
peptide degradation by serum proteases may then reasonably be ques-
DCs pulsed with low concentrations of peptide (46 – 49). There-
tioned. Interestingly, both the serum proteases that degrade tumor Ags
fore, the quality of a peptide vaccine is probably determined not
(Refs. 19 and 24 and this study), those expressed by human DCs (40)
only by the density, but also by the duration, of the presence of
able to degrade synthetic class I peptides (41), and those expressed by
peptide at the injection site for loading onto activated DCs in vivo.
T cells present in the skin and the afferent lymph (42), such as CD13
Thus, peptide protection against protease degradation appears to be
and CD26, display amino- and dipeptidyl-peptidase activities. We
an attractive and effective way to control the concentration and
may thus assume that modified peptides that resist serum proteases
local persistence of an antigenic peptide after injection. In this
probably resist peptidases expressed by DCs and remain stable after
context, the use of peptides protected against degradation should
s.c. injection. Nevertheless, direct determination of the fate and sta-
then allow the optimization of immunization protocols for cancer
bility of antigenic peptides in the interstitial space of the dermis and/or
vaccine. However, concerns may be raised about possible negative
effects that injection of nondegradable peptides may have. In par-
Modifications of the peptide structure leading to resistance to
ticular, it has been reported that due to persistence, diffusion of
proteolysis may alter both the affinity of peptide-MHC binding and
such peptides into the systemic circulation could induce peripheral
the efficiency of CTL recognition. The impact on peptide-MHC
immune tolerance (50). A recent study has also shown pharmaco-
binding and CTL recognition clearly differed among the structural
kinetic differences between a T cell-tolerizing and a T cell-acti-
modifications. As previously described, we show that peptide bond
vating peptide (51). Indeed, knowledge of the pharmacokinetics of
alteration is deleterious for peptide-MHC binding (20, 24, 27). The
peptides is a prerequisite before their possible use in peptide-based
results presented here indicate that amino acid modifications such
immunotherapy. Pharmacokinetic studies of the peptidase-resis-
as ␣-methylation, N-hydroxylation, or -amino acids are among
tant Melan-A peptides are thus currently underway in our labora-
the most efficient to protect against proteolysis and yet cause min-
tories. In addition, the demonstration of increased in vitro immu-
imal reduction of the peptide antigenicity. Interestingly, despite an
nogenicity of the protected peptide should also be extended in vivo
overall decrease in MHC presentation, some of the doubly pro-
and validated. In particular, the correlation between high protease
tected analogs sensitized target cells to lysis by the Melan-A-spe-
resistance and increased immunogenicity remains to be addressed
cific CTL with similar or even higher efficacy than the parental
in vivo. Immunization of transgenic HLA-A2/Kb mice with dif-
peptide. To our knowledge, this is the first time that such an ob-
ferent kinds of adjuvant or several doses of peptide could allow an
A NEW GENERATION OF MELAN-A/MART-1 Ag DERIVATIVES
evaluation of the benefit of the increased resistance to proteolysis
high performance liquid chromatography/electrospray ionization mass spectrom-
on immunogenicity of antigenic peptides. These experiments are
etry: application to the design of peptidase-resistant analogs. Rapid Commun. Mass Spectrom. 12:557.
currently being performed in our laboratories.
20. Brinckerhoff, L. H., V. V. Kalashnikov, L. W. Thompson, G. V. Yamshchikov,
In conclusion, the results shown in the present study demon-
R. A. Pierce, H. S. Galavotti, V. H. Engelhard, and C. L. Slingluff, Jr. 1999. Terminal modifications inhibit proteolytic degradation of an immunogenic
strate that protease-resistant, nonnatural tumor Ag derivatives can
MART-1(27–35) peptide: implications for peptide vaccines. Int. J. Cancer 83:
be highly immunogenic and potent activators of melanoma-spe-
cific CTL. They may represent promising new tools for molecular
21. Valmori, D., J. F. Romero, Y. Men, J. L. Maryanski, P. Romero, and G. Corradin.
1994. Induction of a cytotoxic T cell response by co-injection of a T helper
peptide and a cytotoxic T lymphocyte peptide in incomplete Freund’s adjuvant(IFA): further enhancement by pre-injection of IFA alone. Eur. J. Immunol. 24:Acknowledgments
22. Powell, M. F., T. Stewart, L. J. Otvos, L. Urge, F. Gaeta, A. Sette, T. Arrhenius,
We thank Immanuel Luescher for invaluable advice and support with the
D. Thomson, K. Soda, and S. M. Colon. 1993. Peptide stability in drug devel-
use of tetramers and Pascal Batard and Sophie Millot for technical
opment. II. Effect of single amino acid substitution and glycosylation on peptide
reactivity in human serum. Pharm. Res. 10:1268.
23. Mayer, U., K. Zimmermann, K. Mann, D. Reinhardt, R. Timpl, and R. Nischt. References
1995. Binding properties and protease stability of recombinant human nidogen. Eur. J. Biochem. 227:681.
1. Boon, T., J. C. Cerottini, B. Van den Eynde, P. van der Bruggen, and A. Van Pel.
24. Ayyoub, M., H. Mazarguil, B. Monsarrat, B. Van den Eynde, and J. E. Gairin.
1994. Tumor antigens recognized by T lymphocytes. Annu. Rev. Immunol. 12:
1999. A structure-based approach to designing non-natural peptides that can ac-
tivate anti-melanoma cytotoxic T cells. J. Biol. Chem. 274:10227.
2. Kawakami, Y., and S. A. Rosenberg. 1997. Human tumor antigens recognized by
25. Stemmer, C., A. Quesnel, A. Prevost-Blondel, C. Zimmermann, S. Muller,
J. P. Briand, and H. Pircher. 1999. Protection against lymphocytic choriomenin-
3. Hofbauer, G. F., J. Kamarashev, R. Geertsen, R. Boni, and R. Dummer. 1998.
gitis virus infection induced by a reduced peptide bond analogue of the H-2Db-
Melan A/MART-1 immunoreactivity in formalin-fixed paraffin-embedded pri-
restricted CD8ϩ T cell epitope GP33. J. Biol. Chem. 274:5550.
mary and metastatic melanoma: frequency and distribution. Melanoma Res.
26. Maillere, B., G. Mourier, M. Herve, and A. Menez. 1995. Fine chemical modi-
fications at N- and C-termini enhance peptide presentation to T cells by increas-
4. Kawakami, Y., S. Eliyahu, C. H. Delgado, P. F. Robbins, K. Sakaguchi,
ing the lifespan of both free and MHC-complexed peptides. Mol. Immunol. 32:
E. Appella, J. R. Yannelli, G. J. Adema, T. Miki, and S. A. Rosenberg. 1994.
Identification of a human melanoma antigen recognized by tumor-infiltrating
27. Calbo, S., G. Guichard, P. Bousso, S. Muller, P. Kourilsky, J. P. Briand, and
lymphocytes associated with in vivo tumor rejection. Proc. Natl. Acad. Sci. USA
J. P. Abastado. 1999. Role of peptide backbone in T cell recognition. J. Immunol.
5. Pittet, M. J., D. Valmori, P. R. Dunbar, D. E. Speiser, D. Lienard, F. Lejeune,
28. Hudrisier, D., and J. E. Gairin. 1998. Peptide-major histocompatibility complex
K. Fleischhauer, V. Cerundolo, J. C. Cerottini, and P. Romero. 1999. High fre-
class I complex: from the structural and molecular basis to pharmacological prin-
quencies of naive Melan-A/MART-1-specific CD8ϩ T cells in a large proportion
ciples and therapeutic applications. Curr. Top. Microbiol. Immunol. 232:75.
of human histocompatibility leukocyte antigen (HLA)-A2 individuals. J. Exp.
29. Garcia, K. C., L. Teyton, and I. A. Wilson. 1999. Structural basis of T cell
recognition. Annu. Rev. Immunol. 17:369.
6. Romero, P., N. Gervois, J. Schneider, P. Escobar, D. Valmori, C. Pannetier,
30. Salter, R. D., D. N. Howell, and P. Cresswell. 1985. Genes regulating HLA class
A. Steinle, T. Wolfel, D. Lienard, V. Brichard, et al. 1997. Cytolytic T lympho-
I antigen expression in T-B lymphoblast hybrids. Immunogenetics 21:235.
cyte recognition of the immunodominant HLA-A*0201-restricted Melan-A/MART-1 antigenic peptide in melanoma. J. Immunol. 159:2366.
31. Fehrentz, J. A., and D. Castro. 1983. An efficient synthesis of optically active
7. Dunbar, P. R., C. L. Smith, D. Chao, M. Salio, D. Shepherd, F. Mirza, M. Lipp,
-(t-butyloxycarbonylamino)-aldehydes from ␣-amino acids. Synthesis 8:676.
A. Lanzavecchia, F. Sallusto, A. Evans, et al. 2000. A shift in the phenotype of
32. Bianco, A., C. Zabel, P. Walden, and G. Jung. 1998. N-hydroxy-amide analogues
Melan-A-specific CTL identifies melanoma patients with an active tumor-specific
of MHC-class I peptide ligands with nanomolar binding affinities. J. Pept. Sci.
immune response. J. Immunol. 165:6644.
8. Romero, P., P. R. Dunbar, D. Valmori, M. Pittet, G. S. Ogg, D. Rimoldi,
33. Du¨rr, H., M. Goodman, and J. Gu¨nther. 1992. Retro-inverso amide bonds be-
J. L. Chen, D. Lienard, J. C. Cerottini, and V. Cerundolo. 1998. Ex vivo staining
tween trifunctional amino acids. Angew. Chem. Int. Ed. Engl. 31:785.
of metastatic lymph nodes by class I major histocompatibility complex tetramers
34. Guichard, G., F. Connan, R. Graff, M. Ostankovitch, S. Muller, J. G. Guillet,
reveals high numbers of antigen-experienced tumor-specific cytolytic T lympho-
J. Choppin, and J. P. Briand. 1996. Partially modified retro-inverso pseudopep-
tides as non-natural ligands for the human class I histocompatibility molecule
9. Rivoltini, L., P. Squarcina, D. J. Loftus, C. Castelli, P. Tarsini, A. Mazzocchi,
F. Rini, V. Viggiano, F. Belli, and G. Parmiani. 1999. A superagonist variant of
35. Guichard, G., A. Zerbib, F. A. Le Gal, J. Hoebeke, F. Connan, J. Choppin,
peptide MART1/Melan A27–35 elicits anti-melanoma CD8ϩ T cells with en-
J. P. Briand, and J. G. Guillet. 2000. Melanoma peptide MART-1(27–35) ana-
hanced functional characteristics: implication for more effective immunotherapy.
logues with enhanced binding capacity to the human class I histocompatibility
molecule HLA-A2 by introduction of a -amino acid residue: implications for
10. Valmori, D., J. F. Fonteneau, C. M. Lizana, N. Gervois, D. Lienard, D. Rimoldi,
recognition by tumor-infiltrating lymphocytes. J. Med. Chem. 43:3803.
V. Jongeneel, F. Jotereau, J. C. Cerottini, and P. Romero. 1998. Enhanced gen-
36. Parkhurst, M. R., M. L. Salgaller, S. Southwood, P. F. Robbins, A. Sette,
eration of specific tumor-reactive CTL in vitro by selected Melan-A/MART-1
S. A. Rosenberg, and Y. Kawakami. 1996. Improved induction of melanoma-
immunodominant peptide analogues. J. Immunol. 160:1750.
reactive CTL with peptides from the melanoma antigen glycoprotein 100 mod-
11. Men, Y., I. Miconnet, D. Valmori, D. Rimoldi, J. C. Cerottini, and P. Romero.
ified at HLA-A*0201-binding residues. J. Immunol. 157:2539.
1999. Assessment of immunogenicity of human Melan-A peptide analogues in
37. Powell, M. F., H. Grey, F. Gaeta, A. Sette, and S. Colon. 1992. Peptide stability
HLA-A*0201/Kb transgenic mice. J. Immunol. 162:3566.
in drug development: a comparison of peptide reactivity in different biological
12. Valmori, D., M. J. Pittet, D. Rimoldi, D. Lienard, R. Dunbar, V. Cerundolo,
F. Lejeune, J. C. Cerottini, and P. Romero. 1999. An antigen-targeted approach
38. Araujo, M. C., R. L. Melo, M. H. Cesari, M. A. Juliano, L. Juliano, and
to adoptive transfer therapy of cancer. Cancer Res. 59:2167.
A. K. Carmona. 2000. Peptidase specificity characterization of C- and N-terminal
13. Kugler, A., G. Stuhler, P. Walden, G. Zoller, A. Zobywalski, P. Brossart,
catalytic sites of angiotensin I-converting enzyme. Biochemistry 39:8519.
U. Trefzer, S. Ullrich, C. A. Muller, V. Becker, et al. 2000. Regression of human
39. Calbo, S., G. Guichard, S. Muller, P. Kourilsky, J. P. Briand, and J. P. Abastado.
metastatic renal cell carcinoma after vaccination with tumor cell-dendritic cell
2000. Antitumor vaccination using a major histocompatibility complex (MHC)
class I-restricted pseudopeptide with reduced peptide bond. J. Immunother. 23:
14. Jaeger, E., H. Bernhard, P. Romero, M. Ringhoffer, M. Arand, J. Karbach,
C. Ilsemann, M. Hagedorn, and A. Knuth. 1996. Generation of cytotoxic T-cell
40. Hart, D. N. 1997. Dendritic cells: unique leukocyte populations which control the
responses with synthetic melanoma-associated peptides in vivo: implications for
primary immune response. Blood 90:3245.
tumor vaccines with melanoma-associated antigens. Int. J. Cancer 66:162.
41. Amoscato, A. A., D. A. Prenovitz, and M. T. Lotze. 1998. Rapid extracellular
15. Offringa, R., S. H. van der Burg, F. Ossendorp, R. E. Toes, and C. J. Melief. 2000.
degradation of synthetic class I peptides by human dendritic cells. J. Immunol.
Design and evaluation of antigen-specific vaccination strategies against cancer.
42. Yawalkar, N., R. E. Hunger, W. J. Pichler, L. R. Braathen, and C. U. Brand. 2000.
16. Melief, C. J., R. Offringa, R. E. Toes, and W. M. Kast. 1996. Peptide-based
Human afferent lymph from normal skin contains an increased number of mainly
cancer vaccines. Curr. Opin. Immunol. 8:651.
memory/effector CD4ϩ cells expressing activation, adhesion and co-stimulatory
17. Falo, L. D. J., L. J. Colarusso, B. Benacerraf, and K. L. Rock. 1992. Serum
molecules. Eur. J. Immunol. 30:491.
proteases alter the antigenicity of peptides presented by class I major histocom-
43. Salgaller, M. L., F. M. Marincola, J. N. Cormier, and S. A. Rosenberg. 1996.
patibility complex molecules. Proc. Natl. Acad. Sci. USA 89:8347.
Immunization against epitopes in the human melanoma antigen gp100 following
18. Widmann, C., J. L. Maryanski, P. Romero, and G. Corradin. 1991. Differential
patient immunization with synthetic peptides. Cancer Res. 56:4749.
stability of antigenic MHC class I-restricted synthetic peptides. J. Immunol. 147:
44. Cormier, J. N., M. L. Salgaller, T. Prevette, K. C. Barracchini, L. Rivoltini,
N. P. Restifo, S. A. Rosenberg, and F. M. Marincola. 1997. Enhancement of
19. Ayyoub, M., B. Monsarrat, H. Mazarguil, and J. E. Gairin. 1998. Analysis of the
cellular immunity in melanoma patients immunized with a peptide from MART-
degradation mechanisms of MHC class I-presented tumor antigenic peptides by
1/Melan A. Cancer J. Sci. Am. 3:37.
45. Toes, R. E., R. J. Blom, R. Offringa, W. M. Kast, and C. J. Melief. 1996. En-
48. Alexander-Miller, M. A., G. R. Leggatt, and J. A. Berzofsky. 1996. Selective
hanced tumor outgrowth after peptide vaccination: functional deletion of tumor-
expansion of high- or low-avidity cytotoxic T lymphocytes and efficacy for adop-
specific CTL induced by peptide vaccination can lead to the inability to reject
tive immunotherapy. Proc. Natl. Acad. Sci. USA 93:4102.
49. Bullock, T. N., T. A. Colella, and V. H. Engelhard. 2000. The density of peptides
46. Zeh, H. J., 3rd, D. Perry-Lalley, M. E. Dudley, S. A. Rosenberg, and J. C. Yang.
displayed by dendritic cells affects immune responses to human tyrosinase and
1999. High avidity CTLs for two self-antigens demonstrate superior in vitro and
glycoprotein 100 in HLA-A2 transgenic mice. J. Immunol. 164:2354.
in vivo antitumor efficacy. J. Immunol. 162:989.
50. Aichele, P., K. Brduscha-Riem, R. M. Zinkernagel, H. Hengartner, and
47. Gervois, N., N. Labarriere, S. Le Guiner, M. C. Pandolfino, J. F. Fonteneau,
H. Pircher. 1995. T cell priming versus T cell tolerance induced by synthetic
Y. Guilloux, E. Diez, B. Dreno, and F. Jotereau. 2000. High avidity melanoma-
reactive cytotoxic T lymphocytes are efficiently induced from peripheral blood
51. Weijzen, S., S. C. Meredith, M. P. Velders, A. G. Elmishad, H. Schreiber, and
lymphocytes on stimulation by peptide-pulsed melanoma cells. Clin. Cancer Res.
W. M. Kast. 2001. Pharmacokinetic differences between a T cell-tolerizing and a
T cell-activating peptide. J. Immunol. 166:7151.
Erityisryhmien investointiavustukset ja saman hankkeen korkotukilainat Valmistelija: toimistosihteeri Asta Juvonen, puh. 03-849 3312,Asumisen rahoitus- ja kehittämiskeskus on antanut ohjeet eri tyis ryh-mien investointiavustusten ja saman hankkeen korkotukilainojen ha-ku me net te lys tä vuodel e 2013. Heinolan kaupunki on julistanut ko. avus tuk set ja korkotukilainat haet ta vik si niin, e
A promise to save 100 000 trauma patients In The Lancet , Christopher Murray and colleagues present trauma patients. Please visit the Trauma Promise website the fi ndings of their 2010 Global Burden of Disease to sign up and make the promise. Study, in which they show that injuries cost the global This promise should not be undertaken lightly. Those population some 300 million years of heal
 A NEW GENERATION OF MELAN-A/MART-1 Ag DERIVATIVES
stimulator cells/well in 24-well plate in a total volume of 2 ml of Iscove’s
24-h treatment in serum (Fig. 1A), although it appeared almost
medium supplemented with 10% human serum, Asn, Arg, and Gln (com-
completely degraded after 2 h (Fig. 1B). This observation can be
plete medium) in the presence of IL-7 (10 ng/ml; R&D Systems, Oxon,
explained by a 2- to 3-log difference in the sensitivity of the assays
U.K.), IL-2 (10 U/ml; Glaxo Wellcome, Geneva, Switzerland), and stim-ulating peptide (1 M). Stimulator cells were prepared as follows. Cells
(biological vs physicochemical) as we previously noted (24). It
(2 ϫ 106) derived from the CD8Ϫ population after miniMACS CD8ϩ lym-phocyte isolation were irradiated (3000 rad) and adjusted to the appropriatevolume before addition to the CD8ϩ-enriched responder cell population.
A NEW GENERATION OF MELAN-A/MART-1 Ag DERIVATIVES
stimulator cells/well in 24-well plate in a total volume of 2 ml of Iscove’s
24-h treatment in serum (Fig. 1A), although it appeared almost
medium supplemented with 10% human serum, Asn, Arg, and Gln (com-
completely degraded after 2 h (Fig. 1B). This observation can be
plete medium) in the presence of IL-7 (10 ng/ml; R&D Systems, Oxon,
explained by a 2- to 3-log difference in the sensitivity of the assays
U.K.), IL-2 (10 U/ml; Glaxo Wellcome, Geneva, Switzerland), and stim-ulating peptide (1 M). Stimulator cells were prepared as follows. Cells
(biological vs physicochemical) as we previously noted (24). It
(2 ϫ 106) derived from the CD8Ϫ population after miniMACS CD8ϩ lym-phocyte isolation were irradiated (3000 rad) and adjusted to the appropriatevolume before addition to the CD8ϩ-enriched responder cell population.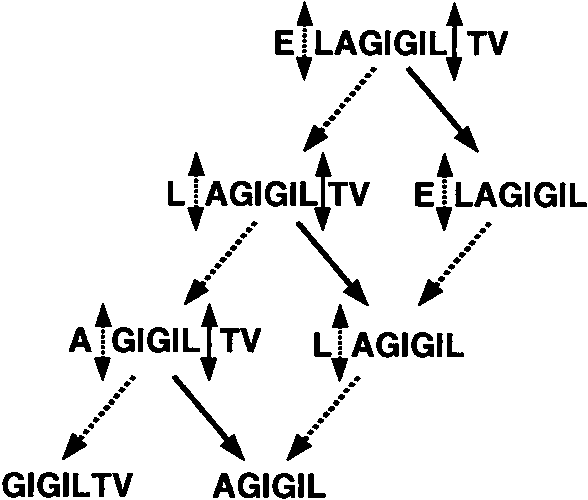 further suggests that few copies of Melan-A
Protection against amino-peptidases was obtained by substitution
able by MS remained in the medium and were still able to stim-
of glutamic acid residue at position 1 (E1) by aa (E1, A1, or
ulate, even though very weakly, the Melan-A-specific CTL.
further suggests that few copies of Melan-A
Protection against amino-peptidases was obtained by substitution
able by MS remained in the medium and were still able to stim-
of glutamic acid residue at position 1 (E1) by aa (E1, A1, or
ulate, even though very weakly, the Melan-A-specific CTL.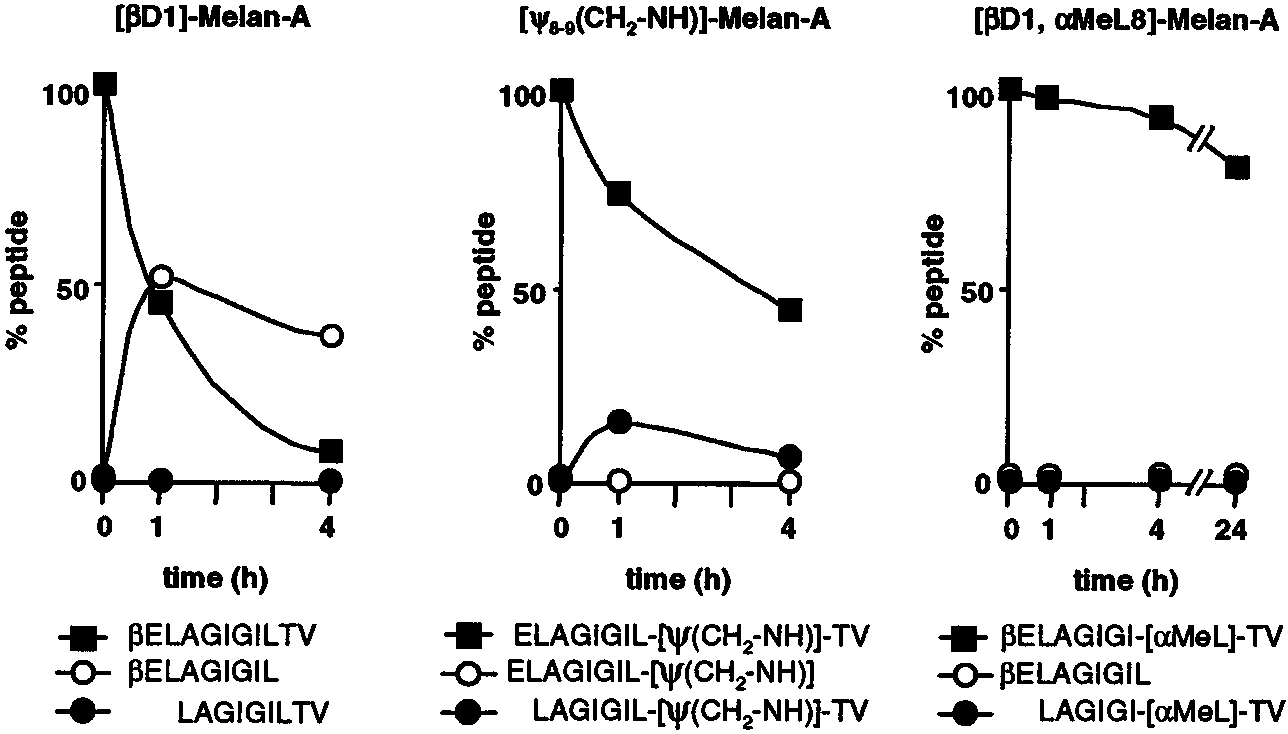
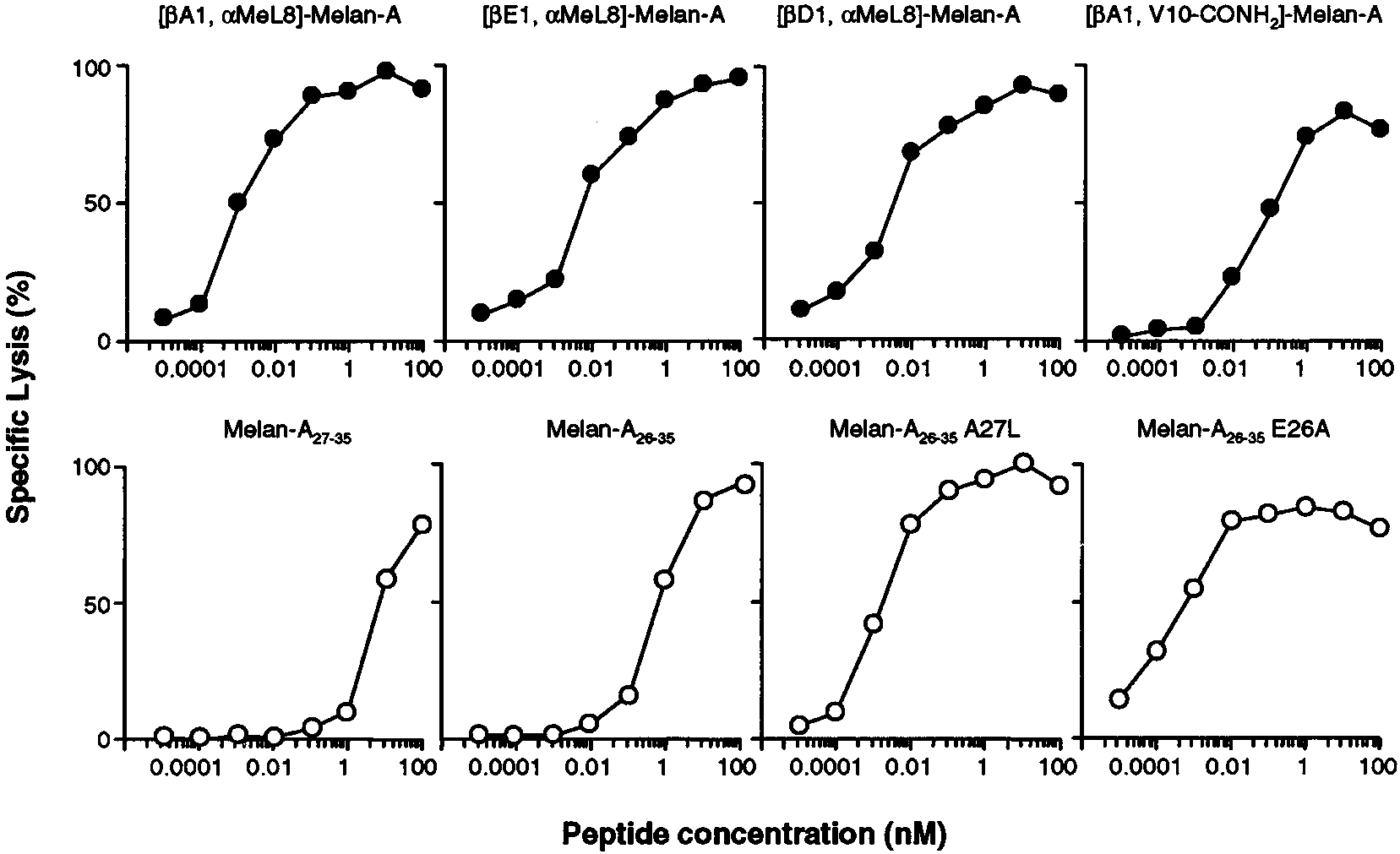 FIGURE 3.
FIGURE 3.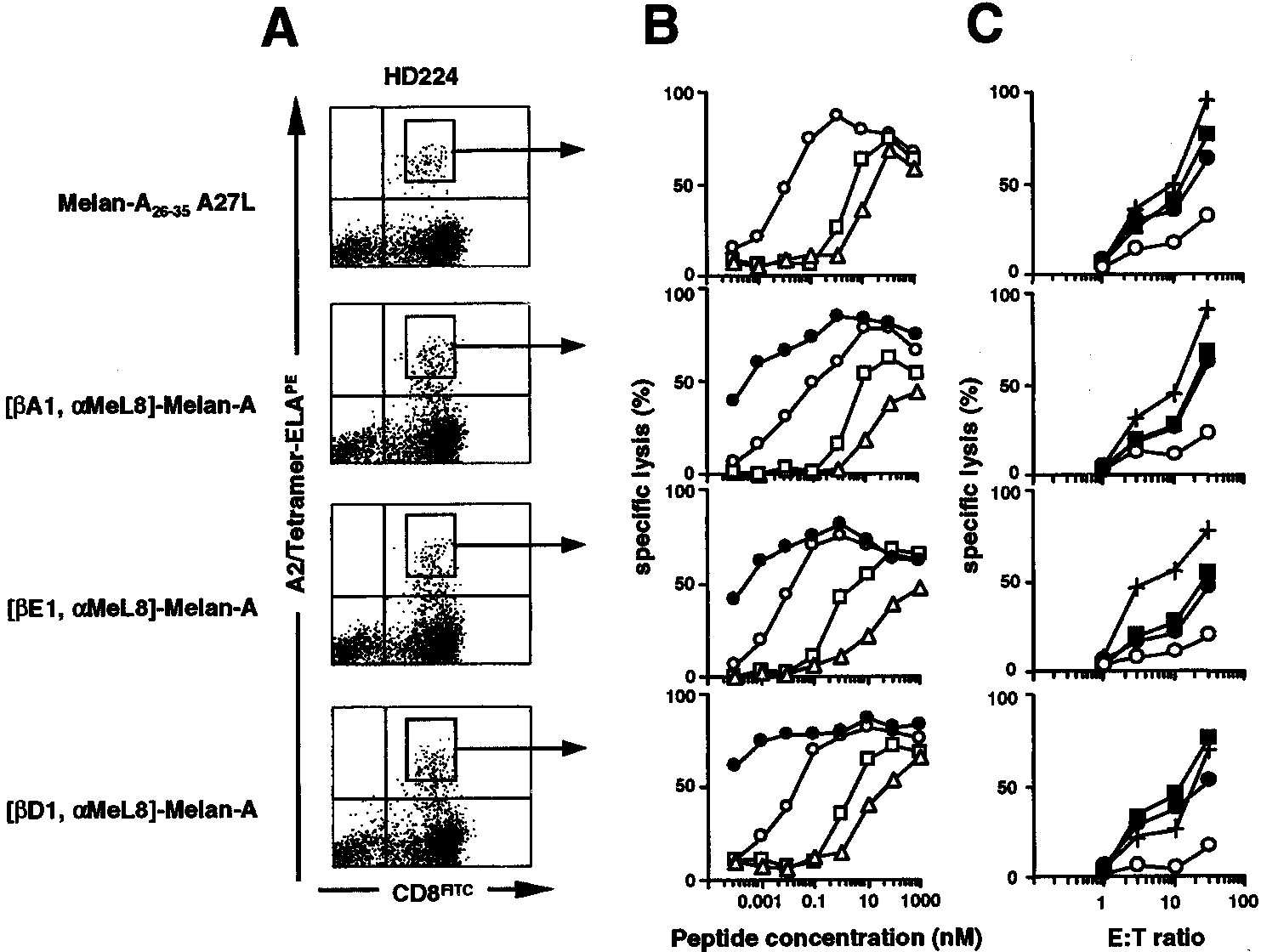 A NEW GENERATION OF MELAN-A/MART-1 Ag DERIVATIVES
Table II. In vitro immunogenicity of Melan-A
less, the use of such immunogenic peptide analogs as a vaccine
may be restricted due to their high susceptibility to protease deg-radation. Thus, the next step was to introduce structural modifica-
tions to obtain proteolysis-resistant antigenic peptides. In the
present study we first elucidated the degradation pathway of the
Melan-A antigenic peptides. Then, according to these results wedesigned structurally modified nonnatural Melan-A analogs that
were potent activators of tumor-reactive CTL.
A NEW GENERATION OF MELAN-A/MART-1 Ag DERIVATIVES
Table II. In vitro immunogenicity of Melan-A
less, the use of such immunogenic peptide analogs as a vaccine
may be restricted due to their high susceptibility to protease deg-radation. Thus, the next step was to introduce structural modifica-
tions to obtain proteolysis-resistant antigenic peptides. In the
present study we first elucidated the degradation pathway of the
Melan-A antigenic peptides. Then, according to these results wedesigned structurally modified nonnatural Melan-A analogs that
were potent activators of tumor-reactive CTL.