La spécificité du tadalafil est liée à sa longue demi-vie, permettant une action qui excède largement celle des autres inhibiteurs de PDE5. L’absorption digestive est complète, avec un pic plasmatique atteint en 2 heures environ. Le métabolisme est réalisé via CYP3A4, produisant des métabolites inactifs éliminés principalement dans les fèces. La sélectivité enzymatique est élevée, réduisant les effets indésirables extra-caverneux. Les réactions indésirables fréquentes incluent céphalées, bouffées vasomotrices et troubles digestifs légers. L’activité pharmacologique est stable, indépendamment de l’ingestion d’aliments. Dans les comparaisons de longue durée, acheter cialis pas cher est mentionné en relation avec les études portant sur la persistance d’efficacité et la constance de la cinétique plasmatique.
Bioinformatica.vil.usal.es
MPMI Vol. 13, No. 12, 2000, pp. 1356–1365. Publication no. M-2000-1002-01R. 2000 The American Phytopathological Society
Restriction Enzyme-Mediated Integration Used to Produce Pathogenicity Mutants of Colletotrichum graminicola M. R. Thon, E. M. Nuckles, and L. J. Vaillancourt
Department of Plant Pathology, S-305 Agricultural Sciences Center-North, University of Kentucky, Lexington
We have developed a restriction enzyme-mediated inser-
pathogenic fungi (Kahmann and Basse 1999; Maier and
tional mutagenesis (REMI) system for the maize pathogen
Shäfer 1999). In a REMI transformation, a restriction enzyme
Colletotrichum graminicola. In this report, we demonstrate
is introduced into the transformation mix. By a process that is
the utility of a REMI-based mutagenesis approach to iden-
poorly understood, the enzyme gains access to the genomic
tify novel pathogenicity genes. Use of REMI increased
DNA in the nucleus of the transformation recipient and intro-
transformation efficiency by as much as 27-fold over
duces double-stranded breaks at its recognition sites. These
transformations with linearized plasmid alone. Ninety-
breaks are recombinogenic with the transforming plasmid
nine transformants were examined by Southern analysis,
DNA that has been linearized with the same restriction en-
and 51% contained simple integrations consisting of one
zyme. In theory, integration of plasmid DNA into a gene at a
copy of the vector integrated at a single site in the genome.
restriction site will cause a mutation, and that mutation will be
All appeared to have a plasmid integration at a unique site. Sequencing across the integration sites of six trans-
The REMI technique is similar to transposon tagging tech-
formants demonstrated that in all cases the plasmid inte-
niques that are available for other organisms and offers some
gration occurred at the corresponding restriction enzyme-
of the same advantages. A restriction enzyme that does not cut
recognition site. We used an in vitro bioassay to identify
within the integrated plasmid can be used to rescue it, along
two pathogenicity mutants among 660 transformants.
with some of the flanking genomic DNA, creating a plasmid
Genomic DNA flanking the plasmid integration sites was
that will replicate in Escherichia coli. The flanking DNA can
used to identify corresponding cosmids in a wild-type
be sequenced and used to isolate cosmids or plasmids from a
genomic library. The pathogenicity of one of the mutants
library, and the rescued plasmid can be introduced back into
was restored when it was transformed with the cosmids.
the recipient organism where homologous recombinationshould recreate the original mutant phenotype, confirming that
Additional keywords: anthracnose leaf blight, anthracnose
the tagged gene is important for a given function.
stalk rot, corn, Glomerella graminicola.
We initiated a REMI mutagenesis project with the goal of
identifying genes that play important roles in establishing
Colletotrichum graminicola (Ces.) G.W. Wils. causes an-
and/or maintaining pathogenic infections of C. graminicola in
thracnose stalk rot and anthracnose leaf blight in maize. These
maize stalks. Using a bioassay for pith colonization and ne-
diseases have become increasingly prevalent in the United
crosis, we screened 660 REMI transformants and identified
States during the last 25 years. Anthracnose stalk rot is the
two strains that had reproducible pathogenicity defects in vi-
most damaging of the two disease phases, and it is considered
tro. These strains also had markedly reduced aggressiveness to
to be one of the most common and economically important of
maize stalks and leaves in vivo. At least one of the mutations
the fungal stalk rots of maize (White 1999). Despite the fact
is likely to have been caused by the integration of the trans-
that C. graminicola is an ubiquitous pathogen with significant
forming plasmid, a conclusion that is based on our ability to
destructive potential (Bergstrom and Nicholson 1999), little is
complement the mutant with cosmid DNA isolated from a
known about the fungal characters that are important for
library by using rescued genomic DNA flanking the integrated
pathogenicity and aggressiveness of C. graminicola on maize
by using plasmid as a probe. This work establishes the feasi-
bility of a REMI approach for isolating pathogenicity mutants
Random mutagenesis studies historically have been effec-
tive for investigations of complex biological processes such aspathogenicity. A transformation technique known as restric-
tion enzyme-mediated integration of plasmid DNA, or REMI,
REMI increased the transformation efficiency
can be used to generate random insertional mutations in plant
of C. graminicola up to 27-fold.
The type of plasmid (circular or linear) and the type and
Corresponding author: L. J. Vaillancourt; Telephone: +1-859-257-2203;
amount of restriction enzyme have been reported to affect the
1356 / Molecular Plant-Microbe Interactions
outcome of REMI transformation (Kahmann and Basse 1999;
ciency. The restriction enzyme quantity needed to achieve the
Maier and Schäfer 1999). We tested the effect of these vari-
optimum gain in transformation efficiency varied depending
ables on the transformation efficiency of C. graminicola with
on which restriction enzyme was used (Table 2). We obtained
plasmid pCB1636, which confers hygromycin resistance (Fig.
the highest transformation efficiencies with 60 units of Hin-
1). Transformation efficiency was typically low (approxi-
mately three transformants per microgram of DNA) when
Twenty-three single-spored transformants were maintained
circular or linearized plasmid was used in the absence of a
on potato dextrose agar (PDA) (Difco Laboratories, Detroit,
restriction enzyme (Table 1). The largest increases were ob-
MI, U.S.A.) without hygromycin for 8 weeks, with transfers
served when a linearized plasmid was used and when the en-
to fresh medium every 2 weeks. All of the transformants re-
zyme used to cut the plasmid was the same as that added to
tained their resistance to hygromycin during the course of this
the transformation mix. When a restriction enzyme other than
that used to digest the plasmid was added, the increase intransformation efficiency was only three- to fivefold. We ob-
Treatment with restriction enzymes had no effect
served a similar small increase in transformation efficiency
on protoplast survival.
when restriction enzymes were used in conjunction with a
Restriction enzymes that produced 3′ overhangs consis-
circular plasmid. We evaluated REMI transformation with
tently resulted in lower transformation efficiencies than en-
eight different restriction enzymes: three that produce 5′ over-
zymes that produced 5′ overhangs (Table 2). We postulated
hangs, three that produce 3′ overhangs, and two that produce
that breaks in the genomic DNA that result in 3′ overhangs
blunt ends. The addition of a restriction enzyme to the trans-
might be less efficiently repaired than other types of DNA
formation mix generally increased the transformation effi-
breaks. We performed an experiment to determine the effectof restriction enzymes alone on protoplast survival. A proce-dure identical to the transformation protocol was used, except
Table 1. Transformation efficiency of Colletotrichum graminicola using different enzyme–plasmid combinations
that no plasmid DNA was included in the transformation mix. Enzyme added to transformation mixa Plasmid treatment HindIII EcoRI BamHI Table 2. Effect of addition of varying quantities of restriction enzymes on transformation efficiency of Colletotrichum graminicola Units of enzyme added to transforma- (transformation efficiency)b Overhang at cleavage site
a For all enzyme treatments, 40 units of enzyme were added to the trans-
b Number of transformants per microgram of DNA.
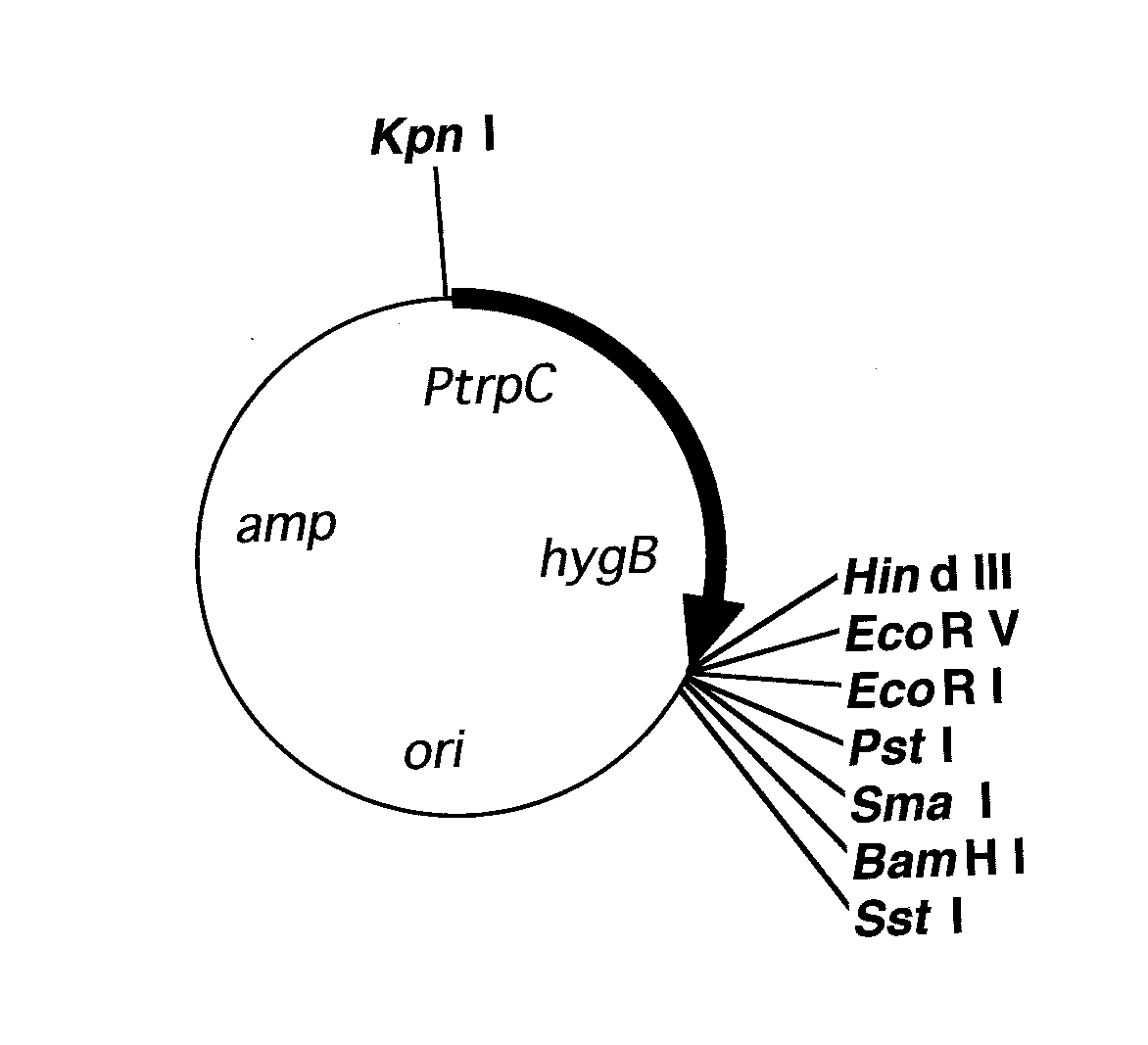
c Numbers in bold represent the averages of two experiments. Fig. 1. Map of plasmid pCB1636 (Sweigard et al. 1997). ori = plasmid
a Plasmid pCB1636 was linearized using the enzyme indicated.
origin of replication. amp = ampicillin resistance gene. PtrpC = Asper-
b Number of transformants per microgram of DNA. gillus nidulans trpC promoter. hygB = bacterial hygromycin B phos-
c Values for three experiments are listed.
d Numbers in bold represent the averages of three experiments.
Vol. 13, No. 12, 2000 / 1357
Serial dilutions of treated protoplasts in polyethylene glycol
ous enzyme combinations (Table 3 and Fig. 2). Of the 99
(PEG) solution were plated in regeneration agar without hy-
transformants, only 18 contained classic REMI integrations in
gromycin. After incubation at room temperature for 3 days,
which a single copy of the plasmid was inserted with the re-
the number of colonies on each dilution plate was recorded.
striction sites present at both plasmid and genomic DNA
We found no significant differences in the number of colonies
junctions (type A) (Fig. 3A). Thirty-two others had single-
after treatment with 0, 20, 40, or 60 units of EcoRI, BamHI or
copy integrations in which one or both of the restriction sites
at the plasmid–genomic DNA junctions had been lost (type B)(Fig. 3A). The remaining 49 transformants had multiple, dis-
More than half of the REMI transformants analyzed
persed integrations; tandem integrations at a single locus;
had single-copy integrations of the plasmid DNA.
some form of rearrangement of the integrated plasmid; or a
We used Southern hybridization analysis to examine 99
combination of these (type C) (Fig. 3A). Each of the 99 trans-
randomly selected transformants that were generated by vari-
formants had a unique restriction pattern, indicating that eachcontained a plasmid integration at a different site. The specificenzyme used for the transformation did not significantly affectthe proportion of type A, B, or C transformants that were re-covered (Table 3). It is notable, however, that none of the 32transformants produced by a linearized plasmid or mis-matched enzymes contained type A integrations. Fungal genomic DNA flanking integrated plasmids could be rescued.
The most desirable integration event for subsequent analy-
sis is to insert one copy of the plasmid at a single, randompoint in the genome (type A or B). Such integrations are com-paratively simple to recover with a restriction enzyme thatdoes not digest the plasmid DNA. Fifty-one percent of thetransformants that we generated were of this type. The remain-der contained complex integrations (type C) that are less easy toanalyze genetically and, reportedly, difficult to rescue (Kahmannand Basse 1999). Typically mutants with complex integrationsare discarded and more tractable mutants are sought. We identi-fied three type C transformants with very interesting pheno-types, however, which we were reluctant to abandon, so weattempted to rescue these more complex integrations.
Southern hybridization analysis indicated that each trans-
formant contained several tandemly arranged copies of thetransforming plasmid at a single locus. Plasmid rescue at-
Fig. 2. Southern hybridization of transformants derived from restriction
tempts using MluI or BglII, enzymes that do not cut within the
enzyme-mediated integration of plasmid DNA using PstI (lanes 1 to 8)
vector, resulted in plasmids that contained complex rear-
or BamHI (lanes 9 to 18). Lanes labeled M contain linearized pCB1636
rangements or deletions of portions of the original transform-
(4.3 kb), which was used as a size marker. Genomic DNA in the upper
ing plasmid. Southern blots indicated that these rearrange-
blot was digested with either PstI (lanes 1 to 8) or BamHI (lanes 9 to18). DNA in the lower blot was digested with KpnI. Both blots were
ments were not present in the genomes of the transformants,
probed with pCB1636. Plasmid integrations were interpreted as follows:
which means they probably occurred during replication of the
type A, 2, 4, 5, 6, 8, 10, 12, 17; type B, 14; type C, 1, 3, 7, 9, 11, 13, 16,
rescued plasmids in E. coli. We then employed a strategy to
18. Transformants 1 and 18 have multiple, dispersed integrations, indi-
recover the two flanking regions separately. Genomic DNA
cated by the presence of multiple bands (left). Transformants 7, 9, and13 have tandem integrations, indicated by the 4.3-kb band (the same size
from each type C transformant was digested with enzymes
as pCB1636) present for these transformants (right).
that contained single recognition sites in the vector (Fig. 3B). Table 3. Plasmid integration events that occurred using various plasmid–restriction enzyme combinations Type of integration Plasmid treatment Enzyme used in transformation Transformants assayed
a Type C transformants are subdivided into those with tandem integrations (Ct) and those with multiple integrations (Cm). 1358 / Molecular Plant-Microbe Interactions
The rescue of DNA from both flanks of the plasmid integra-
type strain and determine whether the integrated plasmid and
tion site was accomplished, in most cases, by digesting the
the mutant phenotype cosegregate. To discern whether an
genomic DNA separately with KpnI or XbaI. The genomic
integrated plasmid DNA segregated normally in a cross, a
DNA was religated and transformed into E. coli. As expected,
randomly selected type A transformant was mated with
a mixture of two plasmid types resulted. One type was derived
M5.001, a compatible wild-type strain, using standard meth-
from internal repeats of the tandemly duplicated plasmid array
ods (Vaillancourt and Hanau 1991). The resulting progeny
within the fungal genome, whereas the other contained flank-
were screened for hygromycin resistance and for the presence
ing genomic DNA (25 to 100% of the plasmids in each ex-
of plasmid pCB1636. Of the 47 progeny assayed, none were
periment contained the desired flanking DNAs).
resistant to hygromycin. Southern analysis demonstrated,however, that three of the 47 contained plasmid DNA, sug-
Plasmid integrations occurred at the recognition site
gesting that these progeny contained nonfunctional forms of
of the restriction enzyme used for REMI.
the hygromycin phosphotransferase gene construct. An un-
We wanted to determine whether REMI integrations typically
linked restriction fragment length polymorphism marker
occurred, as predicted by theory, at the recognition site of the
(Vaillancourt et al. 2000) segregated as expected (1:1) among
restriction enzyme used in the transformation as well as whether
these progeny, suggesting that there was nothing abnormal
deletions of genomic DNA at the plasmid integration sites were
about the cross itself (data not shown).
common. To test these possibilities, we sequenced the genomic
A type C transformant was also crossed with M5.001. Nine of
DNA at the plasmid integration sites in several REMI transfor-
the 15 progeny analyzed were hygromycin resistant, and South-
mants. In addition to the three type C transformants mentioned
ern hybridization confirmed that plasmid DNA was present in
above, we also rescued flanking genomic DNA from one type A
these progeny and absent in the wild-type progeny. Southern
transformant and two type B transformants with either BglII or
blots also revealed that three of the nine hygromycin-resistant
MluI. The plasmid–genomic DNA junctions were sequenced in
progeny contained only a single copy of the plasmid, whereas
each case, and polymerase chain reaction (PCR) primers were
the type C parent and remaining hygromycin-resistant progeny
designed based on the sequence to amplify the corresponding
contained two or more tandemly repeated copies of the plasmid.
region from wild-type (strain M1.001) DNA.
Neither plasmid nor genomic DNA sequences had been de-
leted in the type A transformant. Furthermore, the plasmidintegration occurred at the recognition site of the enzyme usedduring REMI transformation. A 135-bp fragment of DNA wasdeleted from the plasmid at the plasmid–genomic DNA junc-tion of one type B transformant, although the genomic se-quence at the integration site of this transformant was intact. As in the type A transformant, the integration occurred at therecognition site of the enzyme used for transformation. Thesequence of the rescued plasmid from the second type B trans-formant indicated that 6 bp of plasmid DNA had been deleted. The primers that were developed using the sequence of therescued genomic DNA failed to amplify the expected PCRproduct, suggesting that a large deletion of the genomic DNAmay have occurred at the plasmid integration site in this trans-formant. The sequence of the rescued genomic DNA, how-ever, was consistent with an integration event at the recogni-tion site of the enzyme used for transformation.
Similar analyses were performed on the three type C trans-
formants, although in one case cosmid clones from a genomicDNA library were used in place of PCR products from thewild-type DNA. In one type C transformant, a single base pairwas deleted from the genomic DNA at the integration site,whereas in the other two transformants integration occurredwithout any loss of genomic DNA. In all three transformants,plasmid integration events occurred at the recognition sites ofthe enzymes used for transformation. Thus, in all six of the
Fig. 3. Plasmid integration types and plasmid rescue strategies of transfor-
REMI transformants analyzed, integration of the REMI plas-
mants obtained with restriction enzyme-mediated integration. A, Types of
mid occurred at the recognition site of the REMI enzyme. In
plasmid integrations. Note that type C transformants include those withtandemly repeated plasmid copies (shown) as well as multiple dispersed
only one case did there appear to be a significant deletion of
copies (not shown). B, Plasmid rescue strategies. Type A and B transfor-
genomic DNA at the site of plasmid integration.
mants can be rescued by cutting with a single restriction enzyme that hasno recognition site within the plasmid. Flanking DNA from type C trans-
Plasmid integrations in REMI transformants
formants with tandemly repeated plasmid copies can be rescued by cutting
were meiotically unstable.
with each of two restriction enzymes with single recognition sites withinthe plasmid. Bars represent the plasmid DNA; the white part of the bar
One way to discover whether a mutation is tagged in a
represents the portion of the plasmid DNA that is required for replication in
REMI transformant is to cross the transformant with a wild-
Escherichia coli. P = PstI. K = KpnI. Bg = BglII. B = BamHI.
Vol. 13, No. 12, 2000 / 1359 Two pathogenicity mutants were identified
the insertion point was amplified from genomic DNA of wild-
among 660 transformants.
type strain M1.001 and sequenced. The sequence matched that
Our screening strategy uses a rapid assay to identify poten-
of the rescued plasmid, and analysis of the wild-type sequence
tial pathogenicity mutants followed by replicated experiments
confirmed that the integration in 9-4 occurred at a PstI site
to confirm the mutant phenotype. We used the rapid in vitro
with no deletion of the genomic DNA.
assay to identify stalk rot mutants, identifying 41 potential
More than 1 kb of genomic DNA from the region spanning
mutants and subjected each of them to replicated in vitro as-
the integration site in strain 9-4 has been sequenced (GenBank
says. Two transformants (strains 6-2 and 9-4) were consis-
accession number AF264878). Two significant matches were
tently reduced in their ability to colonize or cause necrosis of
found when these sequences were used to search GenBank
maize internode tissues (Fig. 4A to E). Experiments in vivo
and the Saccharomyces genome database with the BLASTX
confirmed the mutant phenotype. In contrast to the wild-type
program and low-complexity sequence filtering. One match
control, neither mutant colonized intact maize stalks and one
occurred with a group of protein-transport proteins that par-
(strain 6-2) caused no detectable stalk discoloration in the in
ticipate in COPII transport vesicle budding from the endo-
vivo experiments (data not shown). Both mutants were tested
plasmic reticulum (E = 7 × 10−28 to 2 × 10−11). The other
for pathogenicity to seedling leaves of maize cultivar Mo940,
match was with cyclophilin proteins from various sources,
an inbred that is highly susceptible to the wild-type strain
including humans, flies, and mice (E = 5 × 10−10 to 5 × 10−6).
M1.001 (Fig. 4F to G). Strain 6-2 caused no symptoms on the
The two regions of similarity are on opposite sides of the
leaves (Fig. 4H), whereas strain 9-4 caused only a few, very
plasmid-insertion site and read in opposite directions. The
predicted amino acid sequence alignments indicate that theplasmid integration occurred within a stretch of approximately
Molecular characterization of strains 6-2 and 9-4.
500 bp between the two putative genes, which could be an
Strain 6-2 was isolated in an experiment in which EcoRI
upstream regulatory region for either gene.
was used as the REMI enzyme. Southern blots of genomic
We conducted an analysis of predicted ORFs and codon us-
DNA from strain 6-2 indicated that this transformant contains
age bias for the rescued sequences with CodonUse. Both of
one complete copy of the plasmid as well as an adjacent in-
the putative genes correspond to ORFs and display significant
complete copy in a reverse orientation. We used NotI-digested
codon bias, suggesting that they may encode functional genes.
genomic DNA to rescue plasmids containing 713 bp of
We also analyzed the sequences surrounding the translation
genomic DNA from one flank of the integration site (plasmid
initiation codons of the two putative genes in 9-4. The start
p62notI.1). Attempts to rescue plasmids containing genomic
position and their context in both cases were similar to those
DNA from the other flank were not successful, perhaps be-
of expressed sequences (Gurr et al. 1987).
cause the copy of the vector adjacent to that flank is incomplete. The genomic DNA from p62notI.1 was used as a probe to re-
Wild-type genomic clones complement the mutation
cover two cosmids from a wild-type genomic library. We then
in strain 6-2.
used one of these cosmids as a template to sequence a 1,309 bp
The rescued genomic DNAs from strains 6-2 and 9-4 were
region that spanned the plasmid integration site (GenBank ac-
used as probes to identify cosmids from a library of the wild-
cession number AF263837). Analysis of the sequence indicated
type strain M1.001 that was constructed in a vector carrying a
that the plasmid integration event occurred at an EcoRI site in
selectable marker for benomyl resistance (J. Rollins and R.
strain 6-2 and that no deletion of genomic DNA occurred.
Hanau, unpublished). One cosmid that hybridized to the
We used the genomic sequence to perform a search of the
probes from strain 9-4 was identified and used to transform
Saccharomyces genome database (available on-line from
protoplasts prepared from strain 9-4. Five benomyl- and hy-
Stanford University, Stanford, CA, U.S.A.) with the BLASTX
gromycin-resistant transformants were tested for pathogenicity
program and low-complexity sequence filtering. Sequence
on maize internode sections in vitro and on leaves in vivo. None
YLR066W was the only database entry to produce high-
of the transformants produced symptoms that were significantly
scoring segment pairs (E = 1.4 × 10−13) and encodes a subunit
different from those caused by strain 9-4 (data not shown).
of a signal peptidase complex. An open reading frame (ORF)
Two overlapping cosmids that hybridized to the probe from
that displays codon usage bias similar to other C. graminicola
strain 6-2 were identified, and each used to transform proto-
genes was detected by the computer program CodonUse (C.
plasts prepared from strain 6-2. Two hygromycin- and benomyl-
Halling, Monsanto Co., St. Louis, MO, U.S.A.). The transla-
resistant transformants were tested for pathogenicity on maize
tion start codon for the ORF begins 776 nucleotides upstream
internode segments in vitro and leaves in vivo. In two separate
of the plasmid integration site and coincides with the start
experiments, the transformants colonized and caused symp-
codon of the yeast signal peptidase subunit.
Strain 9-4 was isolated in an experiment in which PstI was
toms on internode segments that were comparable to those
used as the REMI enzyme and contained a single integration
caused by the wild-type strain (Fig. 4J to K). One transfor-
of two or more tandemly repeated copies of the plasmid.
mant produced foliar lesions that were similar to those caused
Southern hybridization analysis indicated that the PstI sites
by the wild type (Fig. 4M), whereas the second caused symp-
between the tandem copies as well as those at the plasmid–
toms that were consistently less severe than those caused by
genomic DNA junction were intact. The genomic DNA
the wild type yet were still significantly greater than those
flanking the integrated vector on both sides was isolated by
rescuing plasmids derived from either BamHI- or KpnI-
The two 6-2 cosmid transformants were digested with KpnI,
digested genomic DNA. The flanking DNA was sequenced
transferred to nylon membranes, and probed with a genomic
and used to design PCR primers. A 611 bp fragment spanning
DNA fragment immediately adjacent to the integration site of
1360 / Molecular Plant-Microbe Interactions Fig. 4. Inoculation of maize internode segments in vitro and leaves of maize seedlings in vivo with strains of Colletotrichum spp. The internode seg- ments have been cut longitudinally, and the inoculation site is oriented to the upper-center of each segment. A, M1.001, the wild-type strain. Note the triangular area of discoloration immediately below the inoculation wound. B, Water control. C, Sorghum pathogen C. sublineolum. This species is not a pathogen of maize. D, Strain 6-2. E, Strain 9-4. F, M1.001, the wild-type strain. G, Water control. H, Strain 6-2. I, Strain 9-4. J, Strain 6-2 transformed with cosmid 41H6. K, Strain 6-2 transformed with cosmid 40F5. L, Strain 6-2 transformed with cosmid 41H6. M, Strain 6-2 transformed with cosmid 40F5.
Vol. 13, No. 12, 2000 / 1361
pCB1636 in strain 6-2. This probe hybridized to a single 6.2-
transformation of C. graminicola in the absence of restriction
kb band in the wild-type DNA and to a 5.8-kb band in strain
enzymes or with enzymes that did not match the one used to
6-2 (Fig. 5). The cosmid transformants contained both the 5.8-
linearize the plasmid never produced type A transformants. Of
and the 6.2-kb bands, indicating that these strains contain the
the six transformants examined in sufficient detail (one type
disrupted region of DNA present in strain 6-2 as well as a
A, two type B, and three type C), all were the result of plas-mid integration events at the REMI restriction enzyme recog-
wild-type copy of the sequence on the integrated cosmids.
nition site. These observations support the hypothesis that theenzyme in the transformation mix facilitates integration of
DISCUSSION
DNA that is linearized with the same enzyme through the
Although REMI protocols have been described for C.
production of compatible cohesive ends. graminicola (Epstein et al. 1998) and for other filamentous
When we performed REMI transformation experiments in
fungi, detailed analyses of transformation conditions and their
the absence of DNA and selection pressure, we observed no
effect on plasmid integrations have been reported in only a
change in protoplast survival. This suggests that C. gramini-
few cases and, as far as we know, have never been described
cola is able to efficiently repair breaks in the genomic DNA
for C. graminicola. Screening a group of transformants for a
caused by restriction enzymes. Alternatively, it is possible that
loss-of-function phenotype is time consuming, so it is impor-
restriction enzymes in combination with DNA could have a
tant to ensure that plasmid integrations occur in the predicted
negative effect on protoplast survival that restriction enzymes
manner and that mutagenized genomic sequences can be res-
alone do not have. It has been suggested that the presence of
cued and characterized once mutants are identified.
transforming DNA may be required for entry of restriction
The preferred integration event for mutagenesis and plas-
enzymes into the nucleus (Maier and Shäfer 1999).
mid rescue is the insertion of one copy of the plasmid in a
As the number of studies involving REMI increases, it has
single random restriction site of the genome (Kahmann and
become clear that the classical REMI event, in which a single
Basse 1999; Sweigard 1996). Large deletions of genomic
copy of the plasmid integrates at a restriction enzyme recog-
DNA occurring in association with REMI transformation are
nition site leaving both flanking restriction sites intact, does
not uncommon, yet can be a serious problem because the
not occur in every case. In REMI experiments with Magna-
DNA contained in the rescued plasmid may not contain the
porthegrisea, Shi et al. (1995) reported that the occurrence of
gene responsible for the mutation (Maier and Shäfer 1999;
integration events in which the flanking restriction sites re-
Sweigard 1996; Sweigard et al. 1998). Transformation effi-
main intact is up to 72% of transformants with some enzymes.
ciencies are typically increased significantly by REMI. If they
This value included complex transformants with multiple in-
are not, this may indicate that damage caused by the restric-
tegrations (type C). Classical REMI integrations occurred in
tion enzyme is not being repaired efficiently and protoplast
approximately 50% of Ustilagomaydis transformants (Bölker
viability is being compromised. In theory, inefficient DNA
et al. 1995), and in C. graminicola, type A REMI integrations
repair increases the probability that additional mutations will
occurred less frequently: only 28% of the time when the same
be unlinked to plasmid integrations (Maier and Shäfer 1999).
enzyme used for transformation was also used to linearize the
In consideration of these principles, it is important to test
transforming DNA (Table 3). Most integration events resulted
various REMI transformation conditions and determine their
in the loss of one or both flanking restriction sites, usually as a
effects on transformation efficiency and the percentage of the
result of the occurrence of small deletions at the ends of the
preferred integration events produced.
linearized plasmid DNA. These deletions may result from
The addition of restriction enzymes to C. graminicola trans-
nuclease digestion of DNA ends during the transformation
formations increased transformation efficiency up to 27-fold.
process or unequal crossovers during plasmid integration at
A previous study of C. graminicola found only very small
small regions of homology with the ends of the linearized
increases in transformation efficiency (three- to fivefold) with
REMI (Epstein et al. 1998). In this study, however, restriction
Insertional mutagenesis has been used to identify patho-
enzymes were used that did not match the cohesive ends of
genicity genes in several other plant-pathogenic filamentous
the linear plasmid. When we performed transformations in
fungi, but not in C. graminicola. In an assay of more than
this manner, we observed similar small increases in efficiency.
5,000 M. grisea REMI transformants, Sweigard et al. (1998)
Significant increases in transformation efficiency were ob-
identified 27 mutants (0.5%) with a reproducible pathogenic-
served only when the enzyme added to the transformation was
ity defect. Pathogenicity defects occurred in approximately 1
the same as that used to linearize the plasmid. Furthermore,
to 2% of REMI transformants of U. maydis (Bölker et al. 1995). In addition, Sweigard et al. (1998) reported the suc-cessful cloning and sequencing of pathogenicity genes identi-fied in this manner. In this study, we identified two transfor-mants with pathogenicity defects out of 660 transformantsassayed (0.3%).
Because the process of transformation can be mutagenic,
Fig. 5. Southern hybridization of wild-type genomic DNA (lane 1),
untagged mutations may result from the REMI procedure
mutant strain 6-2 (lane 2), strain 6-2 transformed with cosmid 41H6
(Kahmann and Basse 1999; Maier and Shäfer 1999; Sweigard
(lane 3), and strain 6-2 transformed with cosmid 40F5 (lane 4). The
et al. 1998). These include deletions, both large and small, and
DNA in each case was cut with KpnI, which has a single recognition site
chromosomal rearrangements. Therefore, it is important to
within the plasmid pCB1636. A rescued 713-bp fragment of genomic
confirm that mutations obtained from a REMI experiment are
DNA that flanks the plasmid integration site in mutant strain 6-2 wasused as the probe.
associated with integrated DNA. This can be done by con-
1362 / Molecular Plant-Microbe Interactions
ducting sexual crosses; rescuing the integrated plasmid and
U.S.A.). The sexually compatible strain M5.001 was isolated
using it to recreate the original mutation by gene disruption; or
from maize leaves in Brazil (Vaillancourt and Hanau 1994).
complementing the mutation with a wild-type copy of the
Conidia of both strains were stored on silica gel (Tuite 1969)
DNA, spanning the plasmid insertion site. Here we demon-
and in 7% dimethylsulfoxide at −80°C (L. Epstein, personal
strated that the phenotype of mutant strain 6-2 was most likely
communication). Cultures were inoculated from either of these
caused by plasmid integration by complementing it with cos-
frozen stocks and maintained on PDA under continuous fluo-
mids that contained the corresponding region from the wild-
rescent illumination at 25 to 27°C.
type strain. We were unable to complement mutant strain 9-4with a cosmid that was recovered from the library. This may
DNA extraction and purification.
indicate that the REMI integration in this strain is not respon-
Genomic DNA was purified from C. graminicola stationary
sible for the mutant phenotype. Alternatively, it is possible
cultures grown in 10 ml of Difco (Difco Laboratories, Detroit,
that the entire gene sequence(s) required for complementation
MI, U.S.A.) potato dextrose broth in a 60 × 15-mm petri dish
of the mutant is not present on the cosmid.
for 5 days at 25 to 27°C under continuous fluorescent light.
Strain 6-2 is nonpathogenic to maize leaves and stalks in
The mycelial mats were collected with sterile toothpicks,
vivo but grows normally in vegetative culture. Our prelimi-
blotted briefly on sterile paper towels, and then placed in 15-
nary studies have given no indication of any abnormalities in
ml polypropylene centrifuge tubes. The mycelium in each tube
spore germination or appressorial formation. Further work
was frozen, lyophilized, and crushed to a fine powder with a
will focus on characterization of the mutant phenotype and
glass stirring rod. The powdered mycelium was mixed with
analysis of the structure and function of this gene, the first to
1.2 ml of DNA extraction buffer (100 mM Tris, pH 7.5; 0.7 M
be identified as playing a significant role in a fungal stalk rot
NaCl; 10 mM EDTA; and 10 g of CTAB per liter) per tube,
disease of maize. It is especially intriguing that the same gene
and the mixture was incubated at 65°C for 30 min. An equal
product appears to be critical for pathogenicity to stalks and
volume of chloroform was added to each sample, and then the
leaves. Genes that confer resistance to C. graminicola in
sample was mixed gently. The samples were centrifuged at
maize stalks are not necessarily the same as those that confer
3,000 × g for 15 min at room temperature, the upper aqueous
resistance to leaf anthracnose (Zuber et al. 1981); thus, it is
phase was removed to a 1.5-ml Eppendorf tube, and the DNA
not clear whether the same host–pathogen interaction occurs
was precipitated with 0.8 volume of isopropanol. After cen-
trifugation, the pellets were washed with 70% ethanol, air
Our work suggests that the analysis of REMI mutants of C.
dried, and dissolved in 100 µl of Tris-EDTA (TE) buffer. Two
graminicola by classical genetic techniques may be problem-
microliters of RNase A solution (10 mg/ml) was added to each
atic as a result of meiotic instability of the integrated plasmid.
sample, and the samples were stored at 4°C.
Such instability has also been reported by others (Epstein etal. 1998). The primary advantage of insertional mutagenesis
Fungal transformation.
versus other types of mutagenesis techniques is that the muta-
Falcate conidia were harvested from 3- to 4-week-old cul-
tion is likely to be tagged with the transforming DNA. If the
tures of C. graminicola strain M1.001 that had been main-
plasmid and flanking genomic DNA can be rescued together,
tained on PDA. The conidia were washed twice by centrifu-
then the rescued plasmid can be used as a gene-disruption
gation with sterile water and resuspended in 100 ml of Fries’
vector to confirm the source of the mutation (Sweigard 1996).
medium. The conidial culture was incubated at 30°C with
Fifty-one percent of transformants have simple integrations
gentle shaking. After 48 h, the culture contained numerous
that are conducive to plasmid rescue and gene disruption by
oval conidia and very little vegetative growth. The culture was
the rescued plasmid. In other cases, rearrangements of the
filtered through Nytex membrane (Tetko Inc., Briarcliff, NY,
plasmid during transformation or tandem integration of the
U.S.A.), and the filtrate was centrifuged at 2000 × g for 5 min
vector sequences make rescue of the plasmid with both
at room temperature. The pelleted oval conidia were resus-
flanking genomic sequences difficult. We have demonstrated,
pended at 1.5 × 108 conidia per milliliter in a filter-sterilized
however, that short regions flanking the plasmid integration
solution of 0.7 M NaCl and 100 mg of Glucanex (Novo
can be rescued from more complex transformants. While the
Nordisk, Dittengen, Switzerland) per milliliter. The conidial
short flanking sequences cannot be used directly for gene dis-
suspension was incubated at 30°C with gentle shaking for 4 to
ruption, they can be used as probes to identify cosmids in a
5 h until the suspension contained predominantly protoplasts
genomic library for use in complementation and gene disrup-
that were free of cell walls. The protoplasts were recovered by
tion experiments. These methods can substitute for classical
centrifugation in a Beckman GS-6R centrifuge (Beckman
genetic approaches, and so we do not consider meiotic insta-
Instruments, Fullerton, CA, U.S.A.) at 2000 × g for 5 min at
bility of integrated DNA to be a serious impediment to the
4°C and resuspended in 10 ml of STC (1.2 M sorbitol, 10 mM
analysis of C. graminicola REMI-derived mutants. This study
Tris base, and 50 mM CaCl2, pH 7.5). After the protoplast
has also demonstrated the feasibility of the REMI mutagenesis
concentration was determined with a hemacytometer, the
approach for dissection of genetic determinants of patho-
protoplasts were collected by centrifugation and resuspended
genicity of C. graminicola to maize pith and leaf tissues.
in STC at a concentration of 108 protoplasts per milliliter. Theprotoplasts were then frozen in STC at −80°C until use. MATERIALS AND METHODS
Prior to transformation, plasmid pCB1636 was purified
with the Qiagen Plasmid MaxiPrep kit (Qiagen Inc.,
Fungal strains and culture conditions.
Chatsworth, CA, U.S.A.). The plasmid was linearized by di-
The C.graminicola wild-type strain M1.00 was obtained
gestion with a restriction enzyme, and the digestion reactions
from R. Hanau (Purdue University, West Lafayette, IN,
were purified with a Qiaex gel extraction kit (Qiagen Inc.) or
Vol. 13, No. 12, 2000 / 1363
by extracting once with phenol plus chloroform (1:1) and once
Sequencing reactions were performed with a PE Applied
with chloroform, precipitating with 1/10 volume of 3 M so-
Biosystems BigDye Terminator Kit and analyzed on a PE
dium acetate and 2 volumes of ethanol. After drying, the
Applied Biosystems Model 310 Genetic Analyzer.
plasmids were dissolved in TE buffer.
Transformations were performed by combining the proto-
Stalk pathogenicity assays.
plasts with 3 µg of linearized plasmid DNA dissolved in 10 µl
The in vitro bioassay was adapted from one described by
of TE buffer in a 50-ml polypropylene centrifuge tube. The
Nicholson and Warren (1976). Maize inbred Mp305 (Toman
mixture was incubated on ice for 20 min, and then 20 to 60 units
and White 1993) was grown in 10-in. pots in the greenhouse
of restriction enzyme were added to the protoplast mixture im-
in a mixture of 1/2 sterilized field soil and 1/2 Promix
mediately before adding 1 ml of a PEG solution (40% PEG
(Premiere Horticulture Ltd., Rivière-du-Loup, PQ, Canada).
3,350 wt/vol; 0.6 M KCl; 50 mM CaCl2; and 50 mM Tris, pH
Plants were fed daily with a solution of 4.75 g of Miracle-Gro
8). This mixture was incubated at room temperature for 20 min,
18-18-21 formulation for tomatoes (Stern’s Miracle-Gro
combined with 40 ml of regeneration agar (1 M sucrose, 1.25%
Products Inc., Port Washington, NY, U.S.A.) per liter. Plants
casein hydrolysate, 1.25% yeast extract, 1.5% agar, and 250 µg
were harvested at a late vegetative stage (V-11) (Ritchie et al.
of hygromycin B per milliliter), and then poured into two petri
1993), and the first four internodes above the brace roots were
dishes. The dishes were incubated at 30°C, and hygromycin-
recovered for use in the assay. The sheath was removed, and
resistant colonies were transferred to PDA containing 50 µg of
the internodes were cut into 2.5-cm segments. The internode
hygromycin B (PDA + Hyg) per milliliter after 4 to 5 days.
segments were rinsed under cold, running tap water for 1 h
Conidia from these primary transformants were spread onto 2%
and then blotted dry on sterile paper towels. Each internode
water agar containing 50 µg of hygromycin B per milliliter, and
segment was wounded with a sterilized dissecting needle to a
after 24 h, a single hygromycin-resistant germling was recov-
depth of 2 mm. Suspensions of falcate conidia of C. gramini-
ered from each transformant and transferred to PDA + Hyg. cola or C. sublineolum were prepared by collecting conidia
After 2 weeks of incubation, conidia were harvested from the
from 2-week-old cultures, washing them twice with water by
cultures and stored on silica gel at −80°C.
centrifugation, and adjusting the suspension to 5 × 106 conidiaper milliliter of water. A 10-µl drop of conidial suspension
Southern hybridization.
was applied to the wound. C. sublineolum, a stalk rot patho-
Approximately 400 ng of restriction enzyme-digested DNA
gen of sorghum that is nonpathogenic to corn, was used in our
was electrophoresed and transferred to Nytran N membranes
experiments as a negative control. The inoculated internode
(Schleicher & Schuell, Keene, NH, U.S.A.). Probes were hy-
segments were incubated in darkness at 30°C in moist, sterile
bridized to the membranes and detected with a DIG DNA
chambers. After 3 days, they were split longitudinally, with
Labeling and Detection Kit (Boehringer Mannheim, Indian-
the cut centered on the inoculation site. The area of the dis-
apolis, IN, U.S.A.) following the manufacturer’s instructions.
colored region of pith under the inoculation site was meas-
Southern hybridization of the genomic library was performed
ured. A small sample of pith tissue was isolated from an area
by standard techniques (Sambrook et al. 1989) with 32P-
on the opposite side of the stalk segment beyond the discol-
labeled probes prepared with an Oligolabelling Kit (Amer-
ored region. The tissue samples were cultured on PDA plus
sham Pharmacia Biotech, Piscataway, NJ, U.S.A.)
100 µg of ampicillin per milliliter and observed for outgrowth
Plasmid rescue.
of C. graminicola. If C. graminicola grew from the pith sam-
Restriction enzyme-digested genomic DNA was extracted
ple, it was considered to be colonized. Each REMI strain was
once with phenol–chloroform, once with chloroform, and then
tested twice in the preliminary assay. If both replications pro-
precipitated with ethanol. One microgram of digested genomic
duced no significant necrosis, the isolate was retested in three
DNA was recircularized in a 200-µl ligation reaction consist-
separate experiments of five replications each in vitro and in
ing of two units of T4 DNA ligase (Life Technologies,
intact plants in the greenhouse to confirm its phenotype. Iso-
Rockville, MD, U.S.A.) and the manufacturer’s buffer. The
lates were retested in the replicated assays only if they pro-
ligation reaction was performed at 16°C overnight. The reac-
duced necrotic symptoms that were less than 20% of the wild
tion was incubated at 65°C for 10 min and precipitated with 1
volume of sodium acetate and 2 volumes of ethanol, washed
The in vivo stalk assay was similar to the in vitro assay, ex-
with 70% ethanol, and dried. The samples were dissolved in 5
cept that the plants remained intact. The sheath tissue was
µl of TE buffer and electroporated into SURE electroporation-
stripped from the first four internodes above the brace roots.
competent cells (Stratagene, La Jolla, CA, U.S.A.).
The plants were laid on their sides, and a wound was madenear the center of the third internode above the prop roots, as
PCR and DNA sequencing.
was done for the in vitro assay. A 10-µl drop of a conidial
Reactions were performed in 50-µl volumes and consisted
suspension of sterile water, prepared the same way as for the
of 0.5 µM of each primer, 0.2 mM deoxynucleoside triphos-
in vitro assay, was applied to each wound. A ring-shaped sec-
phate, 0.6 units of Taq DNA Polymerase (Life Technologies,
tion of a sterile microfuge tube was placed over each spore
droplet, and the wound site was sealed with Parafilm to create
was supplied with the Taq enzyme. Thermal cycling was per-
a humidity chamber. The inoculated plants were returned to
formed in a PE Applied Biosystems (Foster City, CA, U.S.A.)
their upright positions on the morning following the inocula-
Model 480 Thermal Cycler with 30 cycles at 94°C for 30 s,
tion and incubated on the greenhouse bench for 3 days. The
55°C for 30 s, and 72°C for 30 s. The primer annealing tem-
humidity chambers were left in place for the entire incubation
perature was optimized for each primer pair.
period. At the end of the experiment, internode sections con-
1364 / Molecular Plant-Microbe Interactions
taining the wounds were recovered and treated in a manner
organization of nuclear genes of filamentous fungi. Pages 93-139 in:
Gene Structure in Eukaryotic Organisms. J. R. Kinghorn, ed. IRLPress, Washington, DC.
Kahmann, R., and Basse, C. 1999. REMI (restriction enzyme mediated
Leaf pathogenicity assays.
integration) and its impact on the isolation of pathogenicity genes in
We used two methods for the inoculation of leaves with C.
fungi attacking plants. Eur. J. Plant Pathol. 105:221-229. graminicola. First, falcate conidia were collected from PDA
Maier, F. J., and Shäfer, W. 1999. Mutagenesis via insertional- or re-
plates grown under continuous light for 2 to 3 weeks. Suspen-
striction enzyme-mediated-integration (REMI) as a tool to tag patho-genicity related genes in plant pathogenic fungi. Biol. Chem.
sions of unwashed falcate conidia were prepared in water at a
concentration of 105 conidia per milliliter, and one drop of
Ritchie, S. W., Hanway, J. J., and Benson, G. O. 1993. How a Corn Plant
Tween 20 was added to each 100 ml of suspension. The spores
Develops. Iowa State University of Science and Technology Coop-
were applied to the leaves of V-3 seedlings with a chromatog-
erative Extension Service, Ames, IA, U.S.A.
Sambrook, J., Fritch, E. F., and Maniatis, T. 1989. Molecular Cloning, A
raphy atomizer (Nicholson and Warren 1976). In the second
Laboratory Manual. 2nd ed. Cold Spring Harbor Laboratory Press,
method, spores were applied directly from PDA cultures with
a cotton swab onto the second and third leaves of V-3 seed-
Shi, Z., Christian, D., and Leung, H. 1995. Enhanced transformation in
lings, a technique adapted from Sweigard et al. (1998). In both
Magnaporthegrisea by restriction enzyme mediated integration of
methods, after inoculation the plants were placed in a mist
plasmid DNA. Phytopathology 85:329-333.
Sweigard, J. A. 1996. A REMI primer for filamentous fungi. Int. Soc.
Mol. Plant-Microbe Interact. Rep. Spring:3-5.
Sweigard, J. A., Carroll, A. M., Farrall, L., Chumley, F. G., and Valent,
ACKNOWLEDGMENTS
B. 1998. Magnaporthegrisea pathogenicity genes obtained throughinsertional mutagenesis. Mol. Plant-Microbe Interact. 11:404-412.
We appreciate the excellent technical assistance of D. Brown, J.
Sweigard, J. A., Chumley, F. G., Carroll, A. M., Farrall, L., and Valent,
Takach, and R. Green. We are also grateful to C. Poneleit for allowing us
B. 1997. A series of vectors for fungal transformation. Fungal Genet.
to increase our maize inbred lines in his nursery plots. These studies
were supported by NRI grant 97-35303-4968 from the U.S. Department
Toman, J., Jr., and White, D. G. 1993. Inheritance of resistance to an-
of Agriculture. This is paper number 00-12-157 from the Kentucky
thracnose stalk rot of corn. Phytopathology 83:981-986.
Agricultural Experiment Station, published with permission of the di-
Tuite, J. 1969. Plant Pathological Methods: Fungi and Bacteria. Burgess
Vaillancourt, L. J., and Hanau, R. M. 1991. A method for genetic analy-
sis of Glomerella graminicola (Colletotrichum graminicola) from
LITERATURE CITED
Vaillancourt, L. J., and Hanau, R. M. 1994. Cotransformation and tar-
Bergstrom, G. C., and Nicholson, R. L. 1999. The biology of corn an-
geted gene inactivation in the maize anthracnose fungus Glomerella
thracnose: Knowledge to exploit for improved management. Plant
graminicola. Appl. Environ. Microbiol. 60:3890-3893.
Vaillancourt, L. J., Du, M., Wang, J., Rollins, J., and Hanau, R. 2000.
Bölker, M., Böhnert, H. U., Braun, K. H., Görl, J., and Kahmann, R.
Genetic analysis of cross fertility between two self-sterile strains of
1995. Tagging pathogenicity genes in Ustilagomaydis by restriction
Glomerella graminicola. Mycologia. 92:430-435.
enzyme mediated integration (REMI). Mol. Gen. Genet. 248:547-552.
White, D. G. 1999. Compendium of Corn Diseases. 3rd ed. American
Epstein, L., Lusnak, K., and Kaur, S. 1998. Transformation-mediated
Phytopathological Society, St. Paul, MN, U.S.A.
developmental mutants of Glomerella graminicola (Colletotrichum
Zuber, M. S., Ainsworth, T. C., Blanco, M. H., and Darrah, L. L. 1981. graminicola). Fungal Genet. Biol. 23:189-203.
Effect of anthracnose leaf blight on stalk rind strength and yield in F1
Gurr, S. J., Unkles, S. E., and Kinghorn, J. R. 1987. The structure and
single crosses in maize. Plant Dis. 65:719-722.
Vol. 13, No. 12, 2000 / 1365
Zamzam Paknahad(Ph.D) Postal Address: Academic Qualification : 1986-1989 : B.S, Biology , Shiraz University 1990-1994: M.Sc, Nutrition in Medicine - Shiraz University of Medical Sciences ,Iran 1996-2002: Ph.D, Nutrition, Tabriz University of Medical Sciences, Iran Areas of Particular Expertise: Teaching Areas: Major Research Involvements: Publication: 1
MATERIAL SAFETY DATA SHEET: TERMINANT ANT, TERMITE AND SPIDER SPRAY Hazardous according to criteria of Worksafe Australia Heiniger Home & Garden Care Pty. Ltd. 5 Devon Street, Lonsdale, South Australia, 5160. MATERIAL SAFETY DATA SHEET: TERMINANT ANT, TERMITE AND SPIDER SPRAY Swal owed: Low toxicity. Toxic effects are not expected unless ingestion of large amounts. Aspiration in th
 outcome of REMI transformation (Kahmann and Basse 1999;
ciency. The restriction enzyme quantity needed to achieve the
Maier and Schäfer 1999). We tested the effect of these vari-
optimum gain in transformation efficiency varied depending
ables on the transformation efficiency of C. graminicola with
on which restriction enzyme was used (Table 2). We obtained
plasmid pCB1636, which confers hygromycin resistance (Fig.
outcome of REMI transformation (Kahmann and Basse 1999;
ciency. The restriction enzyme quantity needed to achieve the
Maier and Schäfer 1999). We tested the effect of these vari-
optimum gain in transformation efficiency varied depending
ables on the transformation efficiency of C. graminicola with
on which restriction enzyme was used (Table 2). We obtained
plasmid pCB1636, which confers hygromycin resistance (Fig.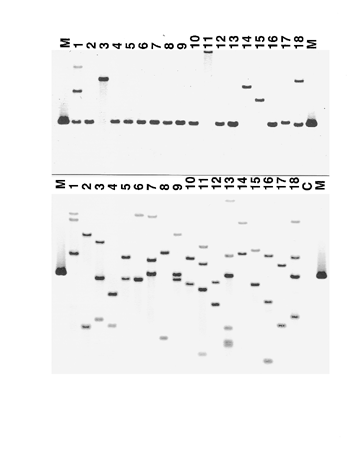 Serial dilutions of treated protoplasts in polyethylene glycol
ous enzyme combinations (Table 3 and Fig. 2). Of the 99
(PEG) solution were plated in regeneration agar without hy-
transformants, only 18 contained classic REMI integrations in
gromycin. After incubation at room temperature for 3 days,
which a single copy of the plasmid was inserted with the re-
the number of colonies on each dilution plate was recorded.
Serial dilutions of treated protoplasts in polyethylene glycol
ous enzyme combinations (Table 3 and Fig. 2). Of the 99
(PEG) solution were plated in regeneration agar without hy-
transformants, only 18 contained classic REMI integrations in
gromycin. After incubation at room temperature for 3 days,
which a single copy of the plasmid was inserted with the re-
the number of colonies on each dilution plate was recorded.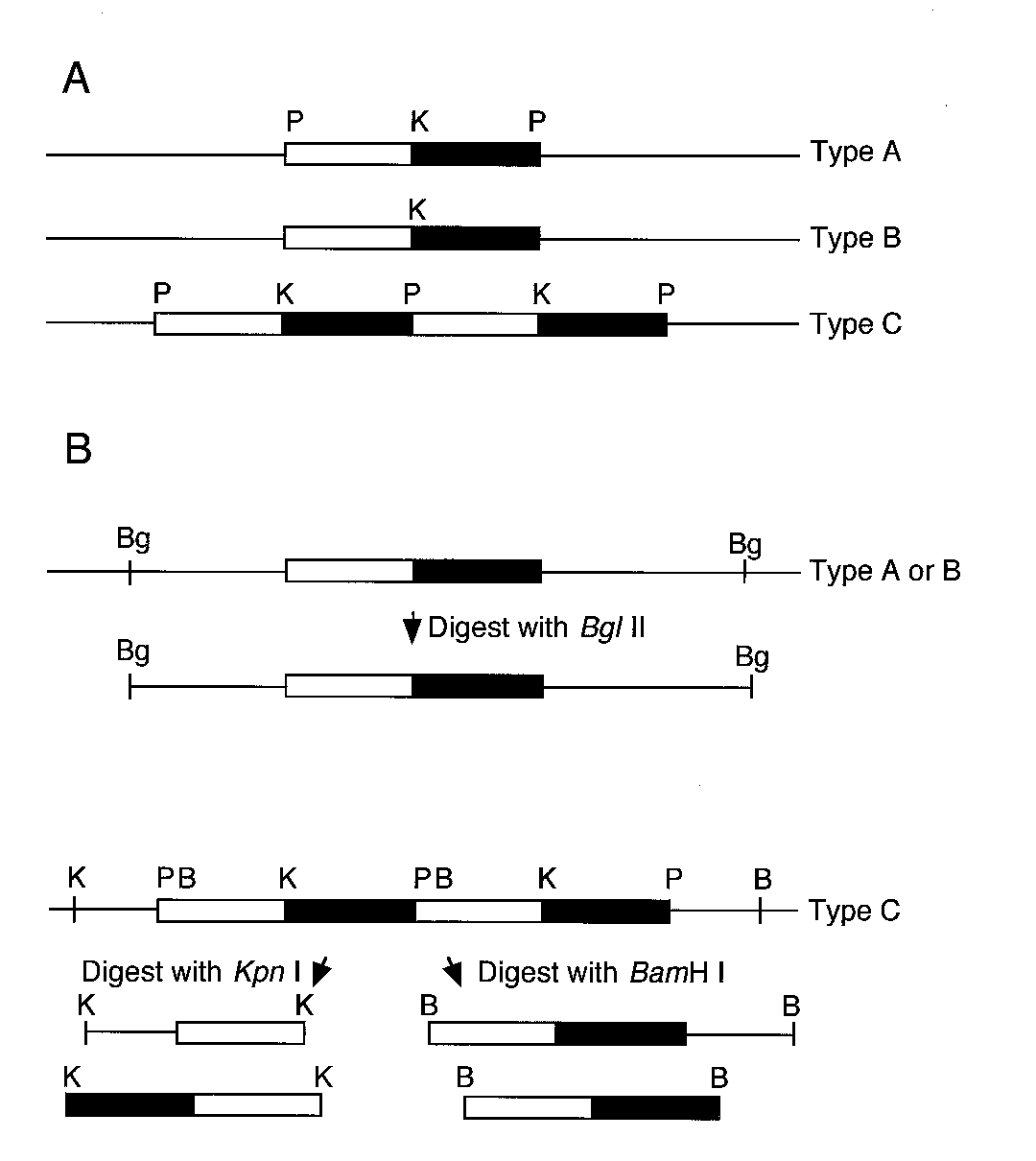 The rescue of DNA from both flanks of the plasmid integra-
type strain and determine whether the integrated plasmid and
tion site was accomplished, in most cases, by digesting the
the mutant phenotype cosegregate. To discern whether an
genomic DNA separately with KpnI or XbaI. The genomic
integrated plasmid DNA segregated normally in a cross, a
DNA was religated and transformed into E. coli. As expected,
randomly selected type A transformant was mated with
a mixture of two plasmid types resulted. One type was derived
M5.001, a compatible wild-type strain, using standard meth-
from internal repeats of the tandemly duplicated plasmid array
ods (Vaillancourt and Hanau 1991). The resulting progeny
within the fungal genome, whereas the other contained flank-
were screened for hygromycin resistance and for the presence
ing genomic DNA (25 to 100% of the plasmids in each ex-
of plasmid pCB1636. Of the 47 progeny assayed, none were
periment contained the desired flanking DNAs).
The rescue of DNA from both flanks of the plasmid integra-
type strain and determine whether the integrated plasmid and
tion site was accomplished, in most cases, by digesting the
the mutant phenotype cosegregate. To discern whether an
genomic DNA separately with KpnI or XbaI. The genomic
integrated plasmid DNA segregated normally in a cross, a
DNA was religated and transformed into E. coli. As expected,
randomly selected type A transformant was mated with
a mixture of two plasmid types resulted. One type was derived
M5.001, a compatible wild-type strain, using standard meth-
from internal repeats of the tandemly duplicated plasmid array
ods (Vaillancourt and Hanau 1991). The resulting progeny
within the fungal genome, whereas the other contained flank-
were screened for hygromycin resistance and for the presence
ing genomic DNA (25 to 100% of the plasmids in each ex-
of plasmid pCB1636. Of the 47 progeny assayed, none were
periment contained the desired flanking DNAs). Fig. 4. Inoculation of maize internode segments in vitro and leaves of maize seedlings in vivo with strains of Colletotrichum spp. The internode seg-
Fig. 4. Inoculation of maize internode segments in vitro and leaves of maize seedlings in vivo with strains of Colletotrichum spp. The internode seg-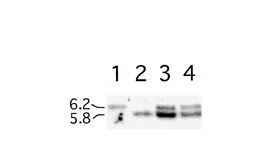 pCB1636 in strain 6-2. This probe hybridized to a single 6.2-
transformation of C. graminicola in the absence of restriction
kb band in the wild-type DNA and to a 5.8-kb band in strain
enzymes or with enzymes that did not match the one used to
6-2 (Fig. 5). The cosmid transformants contained both the 5.8-
linearize the plasmid never produced type A transformants. Of
and the 6.2-kb bands, indicating that these strains contain the
the six transformants examined in sufficient detail (one type
disrupted region of DNA present in strain 6-2 as well as a
A, two type B, and three type C), all were the result of plas-mid integration events at the REMI restriction enzyme recog-
wild-type copy of the sequence on the integrated cosmids.
pCB1636 in strain 6-2. This probe hybridized to a single 6.2-
transformation of C. graminicola in the absence of restriction
kb band in the wild-type DNA and to a 5.8-kb band in strain
enzymes or with enzymes that did not match the one used to
6-2 (Fig. 5). The cosmid transformants contained both the 5.8-
linearize the plasmid never produced type A transformants. Of
and the 6.2-kb bands, indicating that these strains contain the
the six transformants examined in sufficient detail (one type
disrupted region of DNA present in strain 6-2 as well as a
A, two type B, and three type C), all were the result of plas-mid integration events at the REMI restriction enzyme recog-
wild-type copy of the sequence on the integrated cosmids.